Какие структурные компоненты входят в состав днк. Код жизни
Дезоксирибонуклеиновая кислота - полимер, состоит из нуклеотидов.
Нуклеотид ДНК состоит из
- азотистого основания (в ДНК 4 типа: аденин, тимин, цитозин, гуанин)
- моносахара дезоксирибозы
- фосфорной кислоты
Нуклеотиды соединяются между собой прочной ковалентной связью через сахар одного нуклеотида и фосфорную кислоту другого. Получается полинуклеотидная цепь .
Две полинуклеотидные цепи соединяются друг с другом слабыми водородными связями между азотистыми основаниями по правилу комплементарности : напротив аденина всегда стоит тимин, напротив цитозина - гуанин (они подходят друг другу по форме и числу водородных связей - между А и Т две связи, между Ц и Г - 3). Получается двойная цепь ДНК, она скручивается в двойную спираль .
Функция ДНК
ДНК входит в состав хромосом, хранит наследственную информацию (о признаках организма, о первичной структуре белков).
ДНК способна к самоудвоению (репликации, редупликации). Самоудвоение происходит в интерфазе перед делением. После удвоения каждая хромосома состоит из двух хроматид, которые во время будущего деления превратятся в дочерние хромосомы. Благодаря самоудвоению каждая из будущих дочерних клеток получит одинаковую наследственную информацию.
Отличия РНК от ДНК по строению
- рибоза вместо дезоксирибозы
- нет тимина, вместо него урацил
- одноцепочечная
Виды РНК
- информационная (матричная) РНК
- переносит информацию о строении белка из ядра (от ДНК) в цитоплазму (к рибосоме);
- меньше всего в клетке;
- транспортная РНК
- переносит аминокислоты к рибосоме;
- самая маленькая, имеет форму клеверного листа;
- рибосомная РНК
- входит в состав рибосом;
- самая большая по размерам и количеству
Задачи на правило комплементарности
Тимина в ДНК столько же, сколько аденина, остальное (до 100%) приходится на цитозин и гуанин, их тоже поровну. Например: если гуанина 15%, значит цитозина тоже 15%, итого 30%, значит, на аденин и тимин приходится 100-30=70%, следовательно аденина 70/2=35% и тимина тоже 35%
Выберите один, наиболее правильный вариант. Благодаря какому процессу в ходе митоза образуются дочерние клетки с набором хромосом, равным материнскому
1) образования хроматид
2) спирализации хромосом
3) растворения ядерной оболочки
4) деления цитоплазмы
Ответ
Рассмотрите рисунок с изображением фрагмента молекулы биополимера. Определите, (А) что служит ее мономером, (Б) в результате какого процесса увеличивается число этих молекул в клетке, (В) какой принцип лежит в основе ее копирования. Для каждой буквы выберите соответствующий термин из предложенного списка.
1) комплементарность
2) репликация
3) нуклеотид
4) денатурация
5) углевод
6) трансляция
7) транскрипция
Ответ
Все перечисленные ниже признаки, кроме двух, используются для описания изображенной на рисунке молекулы органического вещества. Определите два признака, «выпадающие» из общего списка, и запишите цифры, под которыми они указаны.
1) выполняет ферментативную функцию
2) хранит и передает наследственную информацию
3) состоит из двух нуклеотидных цепей
4) в комплексе с белками образует хромосомы
5) участвует в процессе трансляции
Ответ
Установите соответствие между характеристикой молекулы нуклеиновой кислоты и ее видом: 1) тРНК, 2) ДНК. Запишите цифры 1 и 2 в правильном порядке.
А) состоит из одной полинуклиотидной цепи
Б) транспортирует аминокислоту к рибосоме
В) состоит из 70-80 остатков нуклеотидов
Г) хранит наследственную информацию
Д) способна к репликации
Е) представляет собой спираль
Ответ
НУКЛЕОТИД ИЗ ДРУГОЙ ПАРЫ
1. В ДНК на долю нуклеотидов с тимином приходится 23%. Определите процентное содержание нуклеотидов с гуанином, входящих в состав молекулы. В ответе запишите соответствующее число.
Ответ
2. В ДНК на долю нуклеотидов с цитозином приходится 13%. Определите процентное содержание нуклеотидов с аденином, входящих в состав молекулы. В ответе запишите только соответствующее число.
Ответ
3. В ДНК на долю нуклеотидов с аденином приходится 18%. Определите процентное содержание нуклеотидов с цитозином, входящих в состав молекулы. В ответе запишите только соответствующее число.
Ответ
4. В ДНК на долю нуклеотидов с тимином приходится 36%. Определите процентное содержание нуклеотидов с гуанином, входящих в состав молекулы. В ответе запишите только соответствующее число.
Ответ
5. В ДНК на долю нуклеотидов с тимином приходится 28%. Определите процентное содержание нуклеотидов с гуанином, входящих в состав молекулы. В ответе запишите только соответствующее число.
Ответ
НУКЛЕОТИД ИЗ ЭТОЙ ЖЕ ПАРЫ
1. Фрагмент молекулы ДНК содержит 15% аденина. Сколько тимина в этом фрагменте ДНК? В ответ запишите только число (количество процентов тимина).
Ответ
2. В некоторой молекуле ДНК на долю нуклеотидов с гуанином приходится 28%. Определите процентное содержание нуклеотидов с цитозином, входящих в состав этой молекулы. В ответе запишите только соответствующее число.
Ответ
3. В некоторой молекуле ДНК на долю нуклеотидов с аденином приходится 37%. Определите процентное содержание нуклеотидов с тимином, входящих в состав этой молекулы. В ответе запишите только соответствующее число.
Ответ
НУКЛЕОТИД - СУММА ОДНОЙ ПАРЫ
1. Какое процентное содержание нуклеотидов с аденином и тимином в сумме содержит молекула ДНК, если доля ее нуклеотидов с цитозином составляет 26% от общего числа? В ответе запишите только соответствующее число.
Ответ
2. В ДНК на долю нуклеотидов с цитозином приходится 15%. Определите процентное содержание нуклеотидов с тимином и аденином в сумме, входящих в состав молекулы. В ответе запишите только соответствующее число.
Ответ
СУММА ОДНОЙ ПАРЫ - НУКЛЕОТИД
1. Какой процент составляют нуклеотиды с аденином в молекуле ДНК, если нуклеотиды с гуанином и цитозином вместе составляют 18%? В ответе запишите только соответствующее число.
Ответ
2. В ДНК на долю нуклеотидов с гуанином и цитозином приходится 36%. Определите процентное содержание нуклеотидов с аденином, входящих в состав молекулы. В ответе запишите только соответствующее число.
Ответ
3. В некоторой молекуле ДНК на долю нуклеотидов с аденином и тимином в сумме приходится 26%. Определите процентное содержание нуклеотидов с гуанином, входящих в состав этой молекулы. В ответе запишите только соответствующее число.
Ответ
4. В некоторой молекуле ДНК на долю нуклеотидов с цитозином и гуанином в сумме приходится 42%. Определите процентное содержание нуклеотидов с аденином, входящих в состав этой молекулы. В ответе запишите только соответствующее число.
Ответ
5. В некоторой молекуле ДНК на долю нуклеотидов с аденином и тимином в сумме приходится 54%. Определите процентное содержание нуклеотидов с цитозином, входящих в состав этой молекулы. В ответе запишите только соответствующее число.
Ответ
СУММА РАЗНЫХ ПАР
1. Фрагмент молекулы ДНК содержит 10% тимина. Сколько аденина и гуанина в сумме в этом фрагменте ДНК? В ответ запишите только количество аденина и гуанина в сумме.
Ответ
2. В ДНК на долю нуклеотидов с тимином приходится 35%. Определите процентное содержание нуклеотидов с цитозином и аденином в сумме, входящих в состав молекулы. В ответе запишите только соответствующее число.
Ответ
Выберите три варианта. Чем молекула ДНК отличается от молекулы иРНК?
1) способна самоудваиваться
2) не может самоудваиваться
3) участвует в реакциях матричного типа
4) не может служить матрицей для синтеза других молекул
5) состоит из двух полинуклеотидных нитей, закрученных в спираль
6) является составной частью хромосом
Ответ
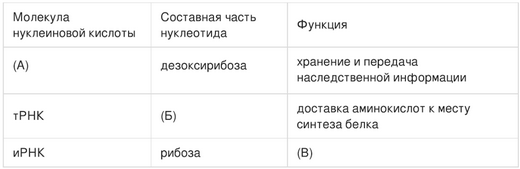
1. Проанализируйте таблицу. Наполните пустые ячейки таблицы, используя понятия и термины, приведенные и списке. Для каждой ячейки, обозначенной буквами, выберите соответствующий термин из предложенного списка.
1) урацил
2) построение тела рибосомы
3) перенос информации о первичной структуре белка
4) рРНК
Ответ

2. Проанализируйте таблицу. Для каждой ячейки, обозначенной буквами, выберите соответствующий термин из предложенного списка.
1) рРНК
2) образование в комплексе с белками тела рибосомы
3) хранение и передача наследственной информации
4) урацил
5) тРНК
6) аминокислота
8) синтез иРНК
Ответ

3. Проанализируйте таблицу «Виды РНК». Для каждой ячейки, обозначенной буквой, выберите соответствующий термин из приведенного списка.
1) мРНК
2) тРНК
3) комплементарна участку молекулы ДНК, несущему информацию о первичной структуре одного белка
4) содержит тимин и дезоксирибозу
5) способна к репликации
6) входит в состав рибосом, участвует в синтезе белка
7) состоит из двух нитей, спирально обвивающих друг друга
Ответ
Выберите один, наиболее правильный вариант. К биологическим полимерам относят молекулу
1) рибозы
2) глюкозы
3) аминокислоты
Ответ
Выберите один, наиболее правильный вариант. Связь, возникающая между азотистыми основаниями двух комплементарных цепей ДНК
1) ионная
2) пептидная
3) водородная
4) ковалентная полярная
Ответ
Выберите один, наиболее правильный вариант. Соединение двух цепей в молекуле ДНК происходит за счет
1) гидрофобных взаимодействий нуклеотидов
2) пептидных связей между азотистыми основаниями
3) взаимодействий комплементарных азотистых оснований
4) ионных взаимодействий нуклеотидов
Ответ
Сколько нуклеотидов с цитозином содержит молекула ДНК, если количество нуклеотидов с тимином 120, что составляет 15% от общего числа? В ответе запишите соответствующее число.
Ответ
В РНК на долю нуклеотидов с урацилом и аденином приходится по 10%. Определите процентное содержание нуклеотидов с тимином входящих в состав комплементарной, двуспиральной цепи ДНК. В ответе запишите только соответствующее число.
Ответ
Участок цепочки ДНК бактериофага лямбда содержит 23 нуклеотида с тимином, сколько нуклеотидов с цитозином в этом участке, если его протяженность 100 нуклеотидов? В ответ запишите только количество нуклеотидов.
Ответ
В молекуле и-РНК содержится 200 нуклеотидов с урацилом, что составляет 10% от общего числа нуклеотидов. Сколько нуклеотидов (в %) с аденином содержит одна из цепей молекулы ДНК? В ответе запишите соответствующее число.
Ответ
Фрагмент молекулы ДНК содержит 60 нуклеотидов. Из них 12 нуклеотидов приходится на тимин. Сколько гуаниновых нуклеотидов содержится в этом фрагменте? В ответе запишите только число.
Ответ
Установите соответствие между признаком нуклеиновой кислоты и ее видом: 1) и-РНК, 2) т-РНК. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) имеет форму клеверного листа
Б) доставляет аминокислоты к рибосоме
В) имеет самые маленькие размеры из нуклеиновых кислот
Г) служит матрицей для синтеза белков
Д) передает наследственную информацию из ядра к рибосоме
Ответ
Установите соответствие между характеристиками и органическими веществами клетки: 1) иРНК, 2) тРНК, 3) рРНК. Запишите цифры 1-3 в порядке, соответствующем буквам.
А) доставляет аминокислоты для трансляции
Б) содержит информацию о первичной структуре полипептида
В) входит в состав рибосом
Г) служит матрицей для трансляции
Д) активизирует аминокислоту
Ответ
1. Все перечисленные ниже признаки, кроме двух, можно использовать для описания молекулы РНК. Определите два признака, «выпадающих» из общего списка, и запишите цифры, под которыми они указаны.
1) состоит из двух полинуклеотидных цепей, закрученных в спираль
2) состоит из одной полинуклеотидной неспирализованной цепи
3) передает наследственную информацию из ядра к рибосоме
4) имеет самые большие размеры из нуклеиновых кислот
5) состоит из нуклеотидов АУГЦ
Ответ
2. Все перечисленные ниже признаки, кроме двух, можно использовать для описания молекулы РНК. Определите два признака, «выпадающих» из общего списка, и запишите цифры, под которыми они указаны.
1) состоит из двух полинуклеотидных цепей, закрученных в спираль
2) переносит информацию к месту синтеза белка
3) в комплексе с белками строит тело рибосомы
4) способна самоудваиваться
5) переносит аминокислоты к месту синтеза белка
Ответ
Выберите один, наиболее правильный вариант. Копией одного или группы генов, несущих информацию о структуре белков, выполняющих одну функцию, является молекула
2) тРНК
3) АТФ
4) иРНК
Ответ
Участок одной из двух цепей молекулы ДНК содержит 300 нуклеотидов с аденином (А), 100 нуклеотидов с тимином (Т), 150 нуклеотидов с гуанином (Г) и 200 нуклеотидов с цитозином (Ц). Сколько нуклеотидов содержится в двух цепях ДНК? Ответ запишите в виде числа.
Ответ
1. Сколько нуклеотидов включает фрагмент двуцепочечной молекулы ДНК, содержащий 14 нуклеотидов с аденином и 20 нуклеотидов с гуанином? В ответе запишите только соответствующее число.
Ответ
2. Сколько нуклеотидов включает в себя фрагмент двуцепочечной молекулы ДНК, если в нём содержится 16 нуклеотидов с тимином и 16 нуклеотидов с цитозином? В ответе запишите только соответствующее число.
Ответ
Все перечисленные ниже признаки, кроме двух, используются для описания изображенной на рисунке схемы строения молекулы органического вещества. Определите два признака, «выпадающие» из общего списка, и запишите цифры, под которыми они указаны.
Молекулярная генетика – раздел генетики, который занимается изучением наследственности на молекулярном уровне.
Нуклеиновые кислоты. Репликация ДНК. Реакции матричного синтеза
Нуклеиновые кислоты (ДНК, РНК) были открыты в 1868 году швейцарским биохимиком И.Ф. Мишером. Нуклеиновые кислоты – линейные биополимеры, состоящие из мономеров – нуклеотидов.
ДНК – структура и функции
Химическую структуру ДНК расшифровали в 1953 г. американский биохимик Дж. Уотсон и английский физик Ф. Крик.
Общая структура ДНК. Молекула ДНК состоит из 2 цепей, которые закручены в спираль (рис. 11) одна вокруг другой и вокруг общей оси. Молекулы ДНК могут содержать от 200 до 2х10 8 пар нуклеотидов. Вдоль спирали молекулы ДНК соседние нуклеотиды располагаются на расстоянии 0,34 нм друг от друга. Полный оборот спирали включает 10 пар нуклеотидов. Его длина составляет 3,4 нм.
Рис . 11 . Схема строения ДНК (двойная спираль)
Полимерность молекулы ДНК. Молекула ДНК – биоплоимер состоит из сложных соединений – нуклеотидов.
Строение нуклеотида ДНК. Нуклеотид ДНК состоит из 3 звеньев: одно из азотистых оснований (аденин, гуанин, цитозин, тимин); дезокисирибоза (моносахарид); остаток фосфорной кислоты (рис. 12).
Различают 2 группы азотистых оснований:
пуриновые – аденин (А), гуанин (Г), содержащие два бензольных кольца;
пиримидиновые – тимин (Т), цитозин (Ц), содержащие одно бензольное кольцо.
В состав ДНК входят следующие виды нуклеотидов: адениновый (А); гуаниновый (Г); цитозиновый (Ц); тиминовый (Т). Названия нуклеотидов соответствуют названиям азотистых оснований, входящих в их состав: адениновый нуклеотид азотистое основание аденин; гуаниновый нуклеотид азотистое основание гуанин; цитозиновый нуклеотид азотистое основание цитозин; тиминовый нуклеотид азотистое основание тимин.
Соединение двух цепей ДНК в одну молекулу
Нуклеотиды А, Г, Ц и Т одной цепи соединены соответственно с нуклеотидами Т, Ц, Г и А другой цепи водородными связями . Между А и Т формируется две водородные связи, а между Г и Ц – три водородные связи (А=Т, Г≡Ц).
Пары оснований (нуклеотидов) А – Т и Г – Ц называют комплементарными, т. е. взаимно соответствующими. Комплементарность – это химическое и морфологическое соответствие нуклеотидов друг другу в парных цепочках ДНК.
5 ’ 3 ’
1 2 3
3’ 5’
Рис. 12 Участок двойной спирали ДНК. Строение нуклеотида (1– остаток фосфорной кислоты; 2– дезоксирибоза; 3– азотистое основание). Соединение нуклеотидов с помощью водородных связей.
Цепи в молекуле ДНК антипараллельны, т. е. направлены в противоположные стороны, так что 3’- конец одной цепи располагается напротив 5’- конца другой цепи. Генетическая информация в ДНК записана в направлении от 5’ конца к 3’ концу. Эта нить называется смысловой ДНК,
поскольку здесь расположены гены. Вторая нить – 3’–5’ служит эталоном хранения генетической информации.
Cоотношение между числом разных оснований в ДНК установлено Э. Чаргаффом в 1949 г. Чаргафф выявил, что у ДНК различных видов количество аденина равно количеству тимина, а количество гуанина – количеству цитозина.
Правило Э. Чаргаффа :
в молекуле ДНК количество A (адениновых) нуклеотидов всегда равно количеству Т (тиминовых) нуклеотидов или отношение ∑ А к ∑ Т=1. Сумма Г (гуаниновых) нуклеотидов равна сумме Ц (цитозиновых) нуклеотидов или отношение ∑ Г к ∑ Ц=1;
сумма пуриновых оснований (А+Г) равна сумме пиримидиновых оснований (Т+Ц) или отношение ∑ (А+Г) к ∑ (Т+Ц)=1;
Способ синтеза ДНК – репликация . Репликация – это процесс самоудвоения молекулы ДНК, осуществляемый в ядре под контролем ферментов. Самоудовоение молекулы ДНК происходит на основе комплементарности – строгого соответствия нуклеотидов друг другу в парных цепочках ДНК. В начале процесса репликации молекула ДНК раскручивается (деспирализуется) на определенном участке (рис. 13), при этом освобождаются водородные связи. На каждой из цепей, образовавшихся после разрыва водородных связей, при участии фермента ДНК-полимиразы, синтезируется дочерняя цепь ДНК. Материалом для синтеза служат свободные нуклеотиды, содержащиеся в цитоплазме клеток. Эти нуклеотиды выстраиваются комплементарно нуклеотидам двух материнских цепей ДНК. Фермент ДНК-полимераза присоединяет комплементарные нуклеотиды к матричной цепи ДНК. Например, к нуклеотиду А матричной цепи полимераза присоединяет нуклеотид Т и, соответственно, к нуклеотиду Г – нуклеотид Ц (рис. 14). Сшивание комплементарных нуклеотидов происходит с помощью фермента ДНК-лигазы . Так путем самоудвоения синтезируются две дочерние цепи ДНК.
Образовавшиеся две молекулы ДНК из одной молекулы ДНК представляют собой полуконсервативную модель , поскольку состоят из старой материнской и новой дочерней цепей и являются точной копией материнской молекулы (рис. 14). Биологический смысл репликации заключается в точной передаче наследственной информации от материнской молекулы к дочерней.

Рис . 13 . Деспирализация молекулы ДНК с помощью фермента
1

Рис . 14 . Репликация – образование двух молекул ДНК из одной молекулы ДНК: 1 – дочерняя молекула ДНК; 2 – материнская (родительская) молекула ДНК.
Фермент ДНК-полимераза может двигаться вдоль цепи ДНК только в направлении 3’ –> 5’. Поскольку комплементарные цепи в молекуле ДНК направлены в противоположные стороны, и фермент ДНК-полимераза может двигаться вдоль цепи ДНК только в направлении 3’–>5’, то и синтез новых цепей идет антипараллельно (по принципу антипараллельности ).
Место локализации ДНК . ДНК содержится в ядре клетки, в матриксе митохондрий и хлоропластов.
Количество ДНК в клетке постоянно и составляет 6,6х10 -12 г.
Функции ДНК:
Хранение и передача в ряду поколений генетической информации молекулам и - РНК;
Структурная. ДНК является структурной основой хромосом (хромосома на 40% состоит из ДНК).
Видоспецифичность ДНК . Нуклеотидный состав ДНК служит критерием вида.
РНК, строение и функции.
Общая структура .
РНК – линейный биополимер, состоящий из одной полинуклеотидной цепи. Различают первичную и вторичную структуры РНК. Первичная структура РНК представляет собой одноцепочечную молекулу, а вторичная структура имеет форму креста и характерна для т- РНК.
Полимерность молекулы РНК . Молекула РНК может включать от 70 нуклеотидов до 30 000 нуклеотидов. Нуклеотиды, входящие в состав РНК, следующие: адениловый (А), гуаниловый (Г), цитидиловый (Ц), урациловый (У). В составе РНК тиминовый нуклеотид замещен на урациловый (У).
Строение нуклеотида РНК.
Нуклеотид РНК включает 3 звена:
азотистое основание (аденин, гуанин, цитозин, урацил);
моносахарид – рибоза (в рибозе присутствует кислород при каждом атоме углерода);
остаток фосфорной кислоты.
Способ синтеза РНК – транскрипция . Транскрипция, как и репликация, – реакция матричного синтеза. Матрицей является молекула ДНК. Реакция протекает по принципу комплементарности на одной из цепей ДНК (рис. 15). Процесс транскрипции начинается с деспирализации молекулы ДНК на определенном участке. На транскрибируемой цепи ДНК имеется промотор – группа нуклеотидов ДНК, с которой начинается синтез молекулы РНК. К промотору присоединяется фермент РНК-полимераза . Фермент активизирует процесс транскрипции. По принципу комплементарности достраиваются нуклеотиды, поступающие из цитоплазмы клетки к транскрибируемой цепи ДНК. РНК-полимераза активизирует выстраивание нуклеотидов в одну цепь и формирование молекулы РНК.
В процессе транскрипции выделяют четыре стадии: 1) связывание РНК-полимеразы с промотором; 2) начало синтеза (инициация); 3) элонгация – рост цепи РНК, т. е. происходит последовательное присоединение нуклеотидов друг к другу; 4) терминация – завершение синтеза и-РНК.

Рис . 15 . Схема транскрипции
1 – молекула ДНК (двойная цепочка); 2 – молекула РНК; 3–кодоны; 4– промотор.
В 1972 г. американские ученые – вирусолог Х.М. Темин и молекулярный биолог Д. Балтимор на вирусах в опухолевых клетках открыли обратную транскрипцию. Обратная транскрипция – переписывание генетической информации с РНК на ДНК. Процесс протекает с помощью фермента обратной транскриптазы .
Виды РНК по функции
Информационная, или матричная РНК (и-РНК, или м-РНК) переносит генетическую информацию с молекулы ДНК к месту синтеза белка – в рибосому. Синтезируется в ядре при участии фермента РНК-полимеразы. Она составляет 5% от всех видов РНК клетки. и- РНК включает от 300 нуклеотидов до 30 000 нуклеотидов (самая длинная цепь среди РНК).
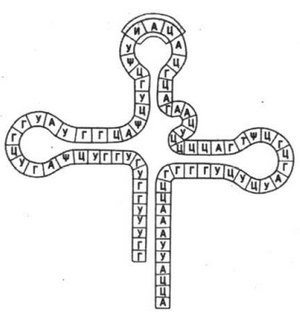
Транспортная РНК (т-РНК) транспортирует аминокислоты к месту синтеза белка,– в рибосому. Имеет форму креста (рис. 16) и состоит из 70 – 85 нуклеотидов. Ее количество в клетке составляет 10-15 % РНК клетки.

Рис. 16. Схема строения т-РНК: А–Г – пары нуклеотидов, соединенные с помощью водородных связей; Д – место прикрепления аминокислоты (акцепторный участок); Е – антикодон.
3. Рибосомная РНК (р-РНК) синтезируется в ядрышке и входит в состав рибосом. Включает примерно 3000 нуклеотидов. Составляет 85% РНК клетки. Этот вид РНК содержатся в ядре, в рибосомах, на эндоплазматической сети, в хромосомах, в матриксе митохондрий, а также в пластидах.
Основы цитологии. Решение типовых задач
Задача 1
Сколько тиминовых и адениновых нуклеотидов содержится в ДНК, если в ней обнаружено 50 цитозиновых нуклеотидов, что составляет 10% от всех нуклеотидов.
Решение. По правилу комплементарности в двойной цепи ДНК цитозин всегда комплемпентарен гуанину. 50 цитозиновых нуклеотидов составляют 10%, следовательно, согласно правилу Чаргаффа, 50 гуаниновых нуклеотидов также составляют 10%, или (если ∑Ц =10%, то и ∑Г =10%).
Сумма пары нуклеотидов Ц + Г равна 20%
Сумма пары нуклеотидов Т + А = 100% – 20 % (Ц + Г) = 80 %
Для того, чтобы узнать, сколько тиминовых и адениновых нуклеотидов содержится в ДНК, нужно составить следующую пропорцию:
50 цитозиновых нуклеотидов → 10 %
Х (Т + А) →80 %
Х = 50х80:10=400 штук
Согласно правилу Чаргаффа ∑А= ∑Т, следовательно ∑А=200 и ∑Т=200.
Ответ: количество тиминовых, как и адениновых нуклеотидов в ДНК, равно 200.
Задача 2
Тиминовые нуклеотиды в ДНК составляют 18% от общего количества нуклеотидов. Определите процент остальных видов нуклеотидов, содержащихся в ДНК.
Решение. ∑Т=18%. Согласно правилу Чаргаффа ∑Т=∑А, следовательно на долю адениновых нуклеотидов также приходится 18 % (∑А=18%).
Сумма пары нуклеотидов Т+А равна 36 % (18 % + 18 % = 36 %). На пару нуклеотидов Ги Ц приходится: Г+Ц=100 % –36 %=64 %. Поскольку гуанин всегда комплементарен цитозину, то их содержание в ДНК будет равным,
т. е. ∑ Г= ∑Ц=32%.
Ответ : содержание гуанина, как и цитозина, составляет 32 %.
Задача 3
20 цитозиновых нуклеотидов ДНК составляют 10% от общего количества нуклеотидов. Сколько адениновых нуклеотидов содержится в молекуле ДНК?
Решение. В двойной цепочке ДНК количество цитозина равно количеству гуанина, следовательно, их сумма составляет: Ц+Г=40 нуклеотидов. Находим общее количество нуклеотидов:
20 цитозиновых нуклеотидов → 10 %
Х (общее количество нуклеотидов) →100 %
Х=20х100:10=200 штук
А+Т=200 – 40=160 штук
Так как аденин комплементарен тимину, то их содержание будет равным,
т. е. 160 штук: 2=80 штук, или ∑А=∑Т=80.
Ответ : в молекуле ДНК содержится 80 адениновых нуклеотидов.
Задача 4
Допишите нуклеотиды правой цепи ДНК, если известны нуклеотиды ее левой цепи: АГА – ТАТ – ГТГ – ТЦТ
Решение. Построение правой цепи ДНК по заданной левой цепи производится по принципу комплементарности – строгого соответствия нуклеотидов друг другу: аденонивый – тиминовый (А–Т), гуаниновый – цитозиновый (Г–Ц). Поэтому нуклеотиды правой цепи ДНК должны быть следующие: ТЦТ – АТА – ЦАЦ – АГА.
Ответ : нуклеотиды правой цепи ДНК: ТЦТ – АТА – ЦАЦ – АГА.
Задача 5
Запишите транскрипцию, если транскрибируемая цепочка ДНК имеет следующий порядок нуклеотидов: АГА – ТАТ – ТГТ – ТЦТ.
Решение . Молекула и-РНК синтезируется по принципу комплеиентарности на одной из цепей молекулы ДНК. Нам известен порядок нуклеотидов в транскрибируемой цепи ДНК. Следовательно, надо построить комплементарную цепь и-РНК. Следует помнить, что вместо тимина в молекулу РНК входит урацил. Следовательно:
Цепь ДНК: АГА – ТАТ – ТГТ – ТЦТ
Цепь и-РНК: УЦУ – АУА –АЦА –АГА.
Ответ : последовательность нуклеотидов и-РНК следующая: УЦУ – АУА – АЦА –АГА.
Задача 6
Запишите обратную транскрипцию, т. е. постройте фрагмент двухцепочечной молекулы ДНК по предложенному фрагменту и-РНК, если цепочка и- РНК имеет следующую последовательность нуклеотидов:
ГЦГ – АЦА – УУУ – УЦГ – ЦГУ – АГУ – АГА
Решение. Обратная транскрипция – это синтез молекулы ДНК на основе генетического кода и-РНК. Кодирующая молекулу ДНК и-РНК имеет следующий порядок нуклеотидов: ГЦГ – АЦА – УУУ – УЦГ – ЦГУ – АГУ – АГА. Комплементарная ей цепочка ДНК: ЦГЦ – ТГТ – ААА – АГЦ – ГЦА – ТЦА – ТЦТ. Вторая цепочка ДНК: ГЦГ–АЦА–ТТТ–ТЦГ–ЦГТ–АГТ–АГА.
Ответ : в результате обратной транскрипции синтезированы две цепочки молекулы ДНК: ЦГЦ – ТГТ – ААА – АГЦ – ГЦА – ТЦА и ГЦГ–АЦА–ТТТ–ТЦГ–ЦГТ–АГТ–АГА.
Генетический код. Биосинтез белка.
Ген – участок молекулы ДНК, содержащий генетическую информацию о первичной структуре одного определенного белка.
Экзон-интронная структура гена эукариот
промотор – участок ДНК (длиной до 100 нуклеотидов), к которому присоединяется фермент РНК-полимераза , необходимый для осуществления транскрипции;
2) регуляторная зона – зона, влияющая на активность гена;
3) структурная часть гена – генетическая информация о первичной структуре белка.
Последовательность нуклеотидов ДНК, несущая генетическую информацию о первичной структуре белка – экзон . Они также входят в состав и-РНК. Последовательность нуклеотидов ДНК, не несущая генетическую информацию о первичной структуре белка – интрон . Они не входят в состав и-РНК. В ходе транскрипции с помощью специальных ферментов происходит вырезание копий интронов из и-РНК и сшивание копий экзонов при образовании молекулы и-РНК (рис. 20). Этот процесс называется сплайсинг .

Рис . 20 . Схема сплайсинга (формирование зрелой и-РНК у эукариот)
Генетический код – система последовательности нуклеотидов в молекуле ДНК, или и-РНК, которая соответствует последовательности аминокислот в полипептидной цепи.
Свойства генетического кода:
Триплетность (АЦА – ГТГ – ГЦГ…)
Генетический код является триплетным, так как каждая из 20 аминокислот кодируется последовательностью трех нуклеотидов (триплетом , кодоном) .
Существует 64 вида триплетов нуклеотидов (4 3 =64).
Однозначность (специфичность)
Генетический код является однозначным, так как каждый отдельный триплет нуклеотидов (кодон) кодирует только одну аминокислоту, или один кодон всегда соответствует одной аминокислоте (таблица 3).
Множественность (избыточность, или вырожденность)
Одна и та же аминокислота может кодироваться несколькими триплетами (от 2 до 6), т. к. белокобразующих аминокислот –20, а триплетов – 64.
Непрерывность
Считывание генетической информации происходит в одном направлении, слева направо. Если произойдет выпадение одного нуклеотида, то при считывании его место займет ближайший нуклеотид из соседнего триплета, что приведет к изменению генетической информации.
Универсальность
Генетический код характерен для всех живых организмов, и одинаковые триплеты кодируют одну и ту же аминокислоту у всех живых организмов.
Имеет стартовые и терминальные триплеты (стартовый триплет – АУГ, терминальные триплеты УАА, УГА, УАГ). Эти виды триплетов не кодируют аминокислоты.
Неперекрываемость (дискретность)
Генетический код является неперекрывающимся, так как один и тот же нуклеотид не может входить одновременно в состав двух соседних триплетов. Нуклеотиды могут принадлежать только одному триплету, а если переставить их в другой триплет, то произойдет изменение генетической информации.
Таблица 3 – Таблица генетического кода
|
Основания кодонов |
|||||
Примечание: сокращенные названия аминокислот даны в соответствии с международной терминологией.
Биосинтез белка
Биосинтез белка – вид пластического обмена веществ в клетке, происходящий в живых организмах под действием ферментов. Биосинтезу белка предшествуют реакции матричного синтеза (репликация – синтез ДНК; транскрипция – синтез РНК; трансляция – сборка молекул белка на рибосомах). В процессе биосинтеза белка выделяют 2 этапа:
транскрипция
трансляция
В ходе транскрипции генетическая информация, заключенная в ДНК, находящейся в хромосомах ядра, передается молекуле РНК. По завершении процесса транскрипции и-РНК выходит в цитоплазму клетки через поры в мембране ядра, располагается между 2 субъединицами рибосомы и участвует в биосинтезе белка.
Трансляция – процесс перевода генетического кода в последовательность аминокислот. Трансляция осуществляется в цитоплазме клетки на рибосомах, которые располагаются на поверхности ЭПС (эндоплазматической сети). Рибосомы – сферические гранулы, диаметром, в среднем, 20 нм, состоящие из большой и малой субъединиц. Молекула и-РНК располагается между двумя субъединицами рибосомы. В процессе трансляции участвуют аминокислоты, АТФ, и-РНК, т-РНК, фермент амино-ацил т-РНК-синтетаза.
Кодон – участок молекулы ДНК, или и-РНК, состоящий из трех последовательно расположенных нуклеотидов, кодирующий одну аминокислоту.
Антикодон – участок молекулы т-РНК, состоящий из трех последовательно расположенных нуклеотидов и комплементарный кодону молекулы и-РНК. Кодоны комплементарны соответствующим антикодонам и соединяются с ними с помощью водородных связей (рис. 21).
Синтез белка начинается со стартового кодона АУГ . От него рибосома
перемещается по молекуле и-РНК, триплет за триплетом. Аминокислоты поступают по генетическому коду. Встраивание их в полипептидную цепь на рибосоме происходит с помощью т-РНК. Первичная структура т-РНК (цепочка) переходит во вторичную структуру, напоминающую по форме крест, и при этом в ней сохраняется комплементарность нуклеотидов. В нижней части т-РНК имеется акцепторный участок, к которому присоединяется аминокислота (рис.16). Активизация аминокислоты осуществляется при помощи фермента аминоацил т-РНК-синтетазы . Суть этого процесса состоит в том, что данный фермент взаимодействует с аминокислотой и с АТФ. При этом формируется тройной комплекс, представленный данным ферментом, аминокислотой и АТФ. Аминокислота обогащается энергией, активизируется, приобретает способность образовывать пептидные связи с соседней аминокислотой. Без процесса активизации аминокислоты полипептидная цепь из аминокислт сформироваться не может.
В противоположной, верхней части молекулы т-РНК содержится триплет нуклеотидов антикодон , с помощью которого т-РНК прикрепляется к комплементарному ему кодону (рис. 22).
Первая молекула т-РНК, с присоединенной к ней активизированной аминокислотой, своим антикодоном прикрепляется к кодону и-РНК, и в рибосоме оказывается одна аминокислота. Затем прикрепляется вторая т-РНК своим антикодоном к соответствующему кодону и-РНК. При этом в рибосоме оказываются уже 2 аминокислоты, между которыми формируется пептидная связь. Первая т-РНК покидает рибосому, как только отдаст аминокислоту в полипептидную цепь на рибосоме. Затем к дипептиду присоединяется 3-я аминокислота, ее приносит третья т-РНК и т. д. Синтез белка останавливается на одном из терминальных кодонов – УАА, УАГ, УГА (рис. 23).

1 – кодон и-РНК; кодоны UCG – УЦГ ; CUA – ЦУА ; CGU – ЦГУ ;
2– антикодон т-РНК; антикодон GAT – ГАТ
Рис . 21 . Фаза трансляции: кодон и-РНК притягивается к антикодону т-РНК соответствующими комплементарными нуклеотидами (основаниями)

Справа крупнейшая спираль ДНК человека, выстроенная из людей на пляже в Варне (Болгария), вошедшая в книгу рекордов Гиннесса 23 апреля 2016 года
Дезоксирибонуклеиновая кислота. Общие сведения
ДНК (дезоксирибонуклеиновая кислота) - своеобразный чертеж жизни, сложный код, в котором заключены данные о наследственной информации. Эта сложная макромолекула способна хранить и передавать наследственную генетическую информацию из поколения в поколение. ДНК определяет такие свойства любого живого организма как наследственность и изменчивость. Закодированная в ней информация задает всю программу развития любого живого организма. Генетически заложенные факторы предопределяют весь ход жизни как человека, так и любого др. организхма. Искусственное или естественное воздействие внешней среды способны лишь в незначительной степени повлиять на общую выраженность отдельных генетических признаков или сказаться на развитии запрограммированных процессов.
Дезоксирибонуклеи́новая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами.
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы (С ) и фосфатной (Ф ) группы (фосфодиэфирные связи).

Рис. 2. Нуклертид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы
В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по винтовой линии.
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином (А-Т ), гуанин — только с цитозином (Г-Ц ). Именно эти пары и составляют «перекладины» винтовой "лестницы" ДНК (см.: рис. 2, 3 и 4).

Рис. 2. Азотистые основания
Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции.

Рис. 3. Репликация ДНК
Расположение базовых комбинаций химических соединений ДНК и количественные соотношения между этими комбинациями обеспечивают кодирование наследственной информации.
Образование новой ДНК (репликация)
- Процесс репликации: раскручивание двойной спирали ДНК — синтез комплементарных цепей ДНК-полимеразой — образование двух молекул ДНК из одной.
- Двойная спираль «расстегивается» на две ветви, когда ферменты разрушают связь между базовыми парами химических соединений.
- Каждая ветвь является элементом новой ДНК. Новые базовые пары соединяются в той же последовательности, что и в родительской ветви.
По завершении дупликации образуются две самостоятельные спирали, созданные из химических соединений родительской ДНК и имеющие с ней одинаковый генетический код. Таким путем ДНК способна перерывать информацию от клетки к клетке.
Более подробная информация:
СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ

Рис. 4 . Азотистые основания: аденин, гуанин, цитозин, тимин
Дезоксирибонуклеиновая кислота (ДНК) относится к нуклеиновым кислотам. Нуклеиновые кислоты - это класс нерегулярных биополимеров, мономерами которых являются нуклеотиды.
НУКЛЕОТИДЫ состоят из азотистого основания , соединенного с пятиуглеродным углеводом (пентозой) - дезоксирибозой (в случае ДНК) или рибозой (в случае РНК), который соединяется с остатком фосфорной кислоты (H 2 PO 3 -).
Азотистые основания бывают двух типов: пиримидиновые основания - урацил (только в РНК), цитозин и тимин, пуриновые основания - аденин и гуанин.

Рис. 5. Структура нуклеотидов (слева), расположение нуклеотида в ДНК (снизу) и типы азотистых оснований (справа): пиримидиновые и пуриновые

Атомы углерода в молекуле пентозы нумеруются числами от 1 до 5. Фосфат соединяется с третьим и пятым атомами углерода. Так нуклеинотиды соединяются в цепь нуклеиновой кислоты. Таким образом, мы можем выделить 3’ и 5’-концы цепи ДНК:

Рис. 6. Выделение 3’ и 5’-концов цепи ДНК
Две цепи ДНК образуют двойную спираль . Эти цепи в спирали сориентированы в противоположных направлениях. В разных цепях ДНК азотистые основания соединены между собой с помощью водородных связей . Аденин всегда соединяется с тимином, а цитозин - с гуанином. Это называется правилом комплементарности .
Правило комплементарности:
| A-T G-C |
Например, если нам дана цепь ДНК, имеющая последовательность
3’- ATGTCCTAGCTGCTCG - 5’,
то вторая ей цепь будет комплементарна и направлена в противоположном направлении - от 5’-конца к 3’-концу:
5’- TACAGGATCGACGAGC- 3’.

Рис. 7. Направленность цепей молекулы ДНК и соединение азотистых оснований с помощью водородных связей
РЕПЛИКАЦИЯ ДНК
Репликация ДНК - это процесс удвоения молекулы ДНК путем матричного синтеза. В большинстве случаев естественной репликации ДНК праймером для синтеза ДНК является короткий фрагмент (создаваемый заново). Такой рибонуклеотидный праймер создается ферментом праймазой (ДНК-праймаза у прокариот, ДНК-полимераза у эукариот), и впоследствии заменяется дезоксирибонуклеотидами полимеразой, выполняющей в норме функции репарации (исправления химических повреждений и разрывов в молекле ДНК).
Репликация происходит по полуконсервативному механизму. Это значит, что двойная спираль ДНК расплетается и на каждой из ее цепей по принципу комплементарности достраивается новая цепь. Дочерняя молекула ДНК, таким образом, содержит в себе одну цепь от материнской молекулы и одну вновь синтезированную. Репликация происходит в направлении от 3’ к 5’ концу материнской цепи.

Рис. 8. Репликация (удвоение) молекулы ДНК
ДНК-синтез - это не такой сложный процесс, как может показаться на первый взгляд. Если подумать, то для начала нужно разобраться, что же такое синтез. Это процесс объединения чего-либо в одно целое. Образование новой молекулы ДНК проходит в несколько этапов:
1) ДНК-топоизомераза, располагаясь перед вилкой репликации, разрезает ДНК для того, чтобы облегчить ее расплетание и раскручивание.
2) ДНК-хеликаза вслед за топоизомеразой влияет на процесс «расплетения» спирали ДНК.
3) ДНК-связывающие белки осуществляют связывание нитей ДНК, а также проводят их стабилизацию, не допуская их прилипания друг к другу.
4) ДНК-полимераза δ
(дельта), согласовано со скоростью движения репликативной вилки, осуществляет синтез
ведущей
цепи
дочерней ДНК в направлении 5"→3" на матрице
материнскойнити ДНК по направлению от ее 3"-конца к 5"-концу (скорость до 100 пар нуклеотидов в секунду). Этим события на данной материнской
нити ДНК ограничиваются.

Рис. 9. Схематическое изображение процесса репликации ДНК: (1) Отстающая цепь (запаздывающая нить), (2) Ведущая цепь (лидирующая нить), (3) ДНК-полимераза α (Polα ), (4) ДНК-лигаза, (5) РНК-праймер, (6) Праймаза, (7) Фрагмент Оказаки, (8) ДНК-полимераза δ (Polδ ), (9) Хеликаза, (10) Однонитевые ДНК-связывающие белки, (11) Топоизомераза.
Далее описан синтез отстающей цепи дочерней ДНК (см. Схему репликативной вилки и функции ферментов репликации)
Нагляднее о репликации ДНК см.
5) Непосредственно сразу после расплетания и стабилизации другой нити материнской молекулы к ней присоединяется ДНК-полимераза α (альфа) и в направлении 5"→3" синтезирует праймер (РНК-затравку) - последовательность РНК на матрице ДНК длиной от 10 до 200 нуклеотидов. После этого фермент удаляется с нити ДНК.
Вместо
ДНК-полимеразы
α
к 3"-концу праймера присоединяется
ДНК-полимераза
ε
.
6)
ДНК-полимераза
ε
(эпсилон)
как бы продолжает удлинять праймер, но в качестве субстрата встраивает
дезоксирибонуклеотиды
(в количестве 150-200 нуклеотидов). В результате образуется цельная нить из двух частей -
РНК
(т.е. праймер) и
ДНК
.
ДНК-полимераза
ε
работает до тех пор, пока не встретит праймер предыдущего
фрагмента Оказаки
(синтезированный чуть ранее). После этого данный фермент удаляется с цепи.
7) ДНК-полимераза β (бета) встает вместо ДНК-полимеразы ε , движется в том же направлении (5"→3") и удаляет рибонуклеотиды праймера, одновременно встраивая дезоксирибонуклеотиды на их место. Фермент работает до полного удаления праймера, т.е. пока на его пути не встанет дезоксирибонуклеотид (еще более ранее синтезированный ДНК-полимеразой ε ). Связать результат свой работы и впереди стоящую ДНК фермент не в состоянии, поэтому он сходит с цепи.
В результате на матрице материнской нити "лежит" фрагмент дочерней ДНК. Он называется
фрагмент Оказаки
.
8) ДНК-лигаза производит сшивку двух соседних фрагментов Оказаки , т.е. 5"-конца отрезка, синтезированного ДНК-полимеразой ε , и 3"-конца цепи, встроенного ДНК-полимеразой β .
СТРОЕНИЕ РНК
Рибонуклеиновая кислота (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Так же, как ДНК, РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом . Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Однако в отличие от ДНК, РНК обычно имеет не две цепи, а одну. Пентоза в РНК представлена рибозой, а не дезоксирибозой (у рибозы присутствует дополнительная гидроксильная группа на втором атоме углевода). Наконец, ДНК отличается от РНК по составу азотистых оснований: вместо тимина (Т ) в РНК представлен урацил (U ) , который также комплементарен аденину.
Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией , то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами - РНК-полимеразами .
Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией, т.е. синтеза белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.

Рис. 10. Отличие ДНК от РНК по азотистому основанию: вместо тимина (Т) в РНК представлен урацил (U), который также комплементарен аденину.
ТРАНСКРИПЦИЯ
Это процесс синтеза РНК на матрице ДНК. ДНК раскручивается на одном из участков. На одной из цепей содержится информация, которую необходимо скопировать на молекулу РНК - эта цепь называется кодирующей. Вторая цепь ДНК, комплементарная кодирующей, называется матричной. В процессе транскрипции на матричной цепи в направлении 3’ - 5’ (по цепи ДНК) синтезируется комплементарная ей цепь РНК. Таким образом, создается РНК-копия кодирующей цепи.
![]()
Рис. 11. Схематическое изображение транскрипции
Например, если нам дана последовательность кодирующей цепи
3’- ATGTCCTAGCTGCTCG - 5’,
то, по правилу комплементарности, матричная цепь будет нести последовательность
5’- TACAGGATCGACGAGC- 3’,
а синтезируемая с нее РНК - последовательность
ТРАНСЛЯЦИЯ
Рассмотрим механизм синтеза белка
на матрице РНК, а также генетический код и его свойства. Также для наглядности по ниже приведенной ссылке рекомендуем посмотреть небольшое видео о процессах транскрипции и трансляции, происходящих в живой клетке:

Рис. 12. Процесс синтеза белка: ДНК кодирует РНК, РНК кодирует белок
ГЕНЕТИЧЕСКИЙ КОД
Генетический код - способ кодирования аминокислотной последовательности белков с помощью последовательности нуклеотидов. Каждая аминокислота кодируется последовательностью из трех нуклеотидов - кодоном или триплетом.
Генетический код, общий для большинства про- и эукариот. В таблице приведены все 64 кодона и указаны соответствующие аминокислоты. Порядок оснований — от 5" к 3" концу мРНК.
Таблица 1. Стандартный генетический код
|
1-е
ние |
2-е основание |
3-е
ние |
|||||||
|
U |
C |
A |
G |
||||||
|
U |
U U U |
(Phe/F) |
U C U |
(Ser/S) |
U A U |
(Tyr/Y) |
U G U |
(Cys/C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(Leu/L) |
U C A |
U A A |
Стоп-кодон** |
U G A |
Стоп-кодон** |
A |
||
|
U U G |
U C G |
U A G |
Стоп-кодон** |
U G G |
(Trp/W) |
G |
|||
|
C |
C U U |
C C U |
(Pro/P) |
C A U |
(His/H) |
C G U |
(Arg/R) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln/Q) |
C GA |
A |
||||
|
C U G |
C C G |
C A G |
C G G |
G |
|||||
|
A |
A U U |
(Ile/I) |
A C U |
(Thr/T) |
A A U |
(Asn/N) |
A G U |
(Ser/S) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys/K) |
A G A |
A |
||||
|
A U G |
(Met/M) |
A C G |
A A G |
A G G |
G |
||||
|
G |
G U U |
(Val/V) |
G C U |
(Ala/A) |
G A U |
(Asp/D) |
G G U |
(Gly/G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
G U A |
G C A |
G A A |
(Glu/E) |
G G A |
A |
||||
|
G U G |
G C G |
G A G |
G G G |
G |
|||||
Среди триплетов есть 4 специальных последовательности, выполняющих функции «знаков препинания»:
- *Триплет AUG , также кодирующий метионин, называется старт-кодоном . С этого кодона начинается синтез молекулы белка. Таким образом, во время синтеза белка, первой аминокислотой в последовательности всегда будет метионин.
- **Триплеты UAA , UAG и UGA называются стоп-кодонами и не кодируют ни одной аминокислоты. На этих последовательностях синтез белка прекращается.
Свойства генетического кода
1. Триплетность . Каждая аминокислота кодируется последовательностью из трех нуклеотидов - триплетом или кодоном.
2. Непрерывность . Между триплетами нет никаких дополнительных нуклеотидов, информация считывается непрерывно.
3. Неперекрываемость . Один нуклеотид не может входить одновременно в два триплета.
4. Однозначность . Один кодон может кодировать только одну аминокислоту.

5. Вырожденность . Одна аминокислота может кодироваться несколькими разными кодонами.

6. Универсальность . Генетический код одинаков для всех живых организмов.
Пример. Нам дана последовательность кодирующей цепи:
3’- CCGATTGCACGTCGATCGTATA - 5’.
Матричная цепь будет иметь последовательность:
5’- GGCTAACGTGCAGCTAGCATAT - 3’.
Теперь «синтезируем» с этой цепи информационную РНК:
3’- CCGAUUGCACGUCGAUCGUAUA - 5’.
Синтез белка идет в направлении 5’ → 3’, следовательно, нам нужно перевернуть последовательность, чтобы «прочитать» генетический код:
5’- AUAUGCUAGCUGCACGUUAGCC - 3’.
Теперь найдем старт-кодон AUG:
5’- AUAUG CUAGCUGCACGUUAGCC - 3’.
Разделим последовательность на триплеты:
звучит следующим образом: информация с ДНК передается на РНК (транскрипция), с РНК - на белок (трансляция). ДНК также может удваиваться путем репликации, и также возможен процесс обратной транскрипции, когда по матрице РНК синтезируется ДНК, но такой процесс в основном характерен для вирусов.

Рис. 13. Центральная догма молекулярной биологии
ГЕНОМ: ГЕНЫ и ХРОМОСОМЫ
(общие понятия)
Геном - совокупность всех генов организма; его полный хромосомный набор.
Термин "геном" был предложен Г. Винклером в 1920 г. для описания совокупности генов, заключенных в гаплоидном наборе хромосом организмов одного биологического вида. Первоначальный смысл этого термина указывал на то, что понятие генома в отличие от генотипа является генетической характеристикой вида в целом, а не отдельной особи. С развитием молекулярной генетики значение данного термина изменилось. Известно, что ДНК, которая является носителем генетической информации у большинства организмов и, следовательно, составляет основу генома, включает в себя не только гены в современном смысле этого слова. Большая часть ДНК эукариотических клеток представлена некодирующими ("избыточными") последовательностями нуклеотидов, которые не заключают в себе информации о белках и нуклеиновых кислотах. Таким образом, основную часть генома любого организма составляет вся ДНК его гаплоидного набора хромосом.
Гены — это участки молекул ДНК, кодирующие полипептиды и молекулы РНК
За последнее столетие наше представление о генах существенно изменилось. Ранее геном называли участок хромосомы, кодирующий или определяющий один признак или фенотипическое (видимое) свойство, например цвет глаз.
В 1940 г. Джордж Бидл и Эдвард Тейтем предложили молекулярное определение гена. Ученые обрабатывали споры гриба Neurospora crassa рентгеновским излучением и другими агентами, вызывающими изменения в последовательности ДНК (мутации ), и обнаружили мутантные штаммы гриба, утратившие некоторые специфические ферменты, что в некоторых случаях приводило к нарушению целого метаболического пути. Бидл и Тейтем пришли к выводу, что ген — это участок генетического материала, который определяет или кодирует один фермент. Так появилась гипотеза «один ген — один фермент» . Позднее эта концепция была расширена до определения «один ген — один полипептид» , поскольку многие гены кодируют белки, не являющиеся ферментами, а полипептид может оказаться субъединицей сложного белкового комплекса.
На рис. 14 показана схема того, как триплеты нуклеотидов в ДНК определяют полипептид - аминокислотную последовательность белка при посредничестве мРНК. Одна из цепей ДНК играет роль матрицы для синтеза мРНК, нуклеотидные триплеты (кодоны) которой комплементарны триплетам ДНК. У некоторых бактерий и многих эукариот кодирующие последовательности прерываются некодирующими участками(так называемыми интронами ).
Современное биохимическое определение гена еще более конкретно. Генами называются все участки ДНК, кодирующие первичную последовательность конечных продуктов, к которым относятся полипептиды или РНК, обладающие структурной или каталитической функцией.
Наряду с генами ДНК содержит и другие последовательности, выполняющие исключительно регуляторную функцию. Регуляторные последовательности могут обозначать начало или конец генов, влиять на транскрипцию или указывать место инициации репликации или рекомбинации. Некоторые гены могут экспрессироваться разными путями, при этом один и тот же участок ДНК служит матрицей для образования разных продуктов.
Мы можем приблизительно рассчитать минимальный размер гена , кодирующего средний белок. Каждая аминокислота в полипептидной цепи кодируется последовательностью из трех нуклеотидов; последовательности этих триплетов (кодонов) соответствуют цепочке аминокислот в полипептиде, который кодируется данным геном. Полипептидная цепь из 350 аминокислотных остатков (цепь средней длины) соответствует последовательности из 1050 п.н. (пар нуклеотидов ). Однако многие гены эукариот и некоторые гены прокариот прерываются сегментами ДНК, не несущими информации о белке, и поэтому оказываются значительно длиннее, чем показывает простой расчет.
Сколько генов в одной хромосоме?
 Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
Как известно, бактериальные клетки имеют хромосому в виде нити ДНК, уложенной в компактную структуру - нуклеоид. Хромосома прокариота Escherichia coli , чей геном полностью расшифрован, представляет собой кольцевую молекулу ДНК (на самом деле, это не правильный круг, а скорее петля без начала и конца), состоящую из 4 639 675 п.н. В этой последовательности содержится примерно 4300 генов белков и еще 157 генов стабильных молекул РНК. В геноме человека примерно 3,1 млрд пар нуклеотидов, соответствующих почти 29 000 генам, расположенным на 24 разных хромосомах.
Прокариоты (Бактерии).
 Бактерия E. coli
имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli
приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами
(рис. 16).
Бактерия E. coli
имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli
приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами
(рис. 16).
Большинство плазмид состоит всего из нескольких тысяч пар нуклеотидов, некоторые содержат более 10000 п. н. Они несут генетическую информацию и реплицируются с образованием дочерних плазмид, которые попадают в дочерние клетки в процессе деления родительской клетки. Плазмиды обнаружены не только в бактериях, но также в дрожжах и других грибах. Во многих случаях плазмиды не дают никаких преимуществ клеткам-хозяевам, и их единственная задача — независимое воспроизведение. Однако некоторые плазмиды несут полезные для хозяина гены. Например, содержащиеся в плазмидах гены могут придавать клеткам бактерий устойчивость к антибактериальным агентам. Плазмиды, несущие ген β-лактамазы, обеспечивают устойчивость к β-лактамным антибиотикам, таким как пенициллин и амоксициллин. Плазмиды могут переходить от клеток, устойчивых к антибиотикам, к другим клеткам того же или другого вида бактерий, в результате чего эти клетки также становятся резистентными. Интенсивное применение антибиотиков является мощным селективным фактором, способствующим распространению плазмид, кодирующих устойчивость к антибиотикам (а также транспозонов, которые кодируют аналогичные гены) среди болезнетворных бактерий, и приводит к появлению бактериальных штаммов с устойчивостью к нескольким антибиотикам. Врачи начинают понимать опасность широкого использования антибиотиков и назначают их только в случае острой необходимости. По аналогичным причинам ограничивается широкое использование антибиотиков для лечения сельскохозяйственных животных.
См. также: Равин Н.В., Шестаков С.В. Геном прокариот // Вавиловский журнал генетики и селекции, 2013. Т. 17. № 4/2. С. 972-984.
Эукариоты.
Таблица 2. ДНК, гены и хромосомы некоторых организмов
|
Общая ДНК, п.н. |
Число хромосом* |
Примерное число генов |
|
|
Escherichia coli (бактерия) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae (дрожжи) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans (нематода) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana (растение) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster (плодовая мушка) |
120 367 260 |
20 000 |
|
|
Oryza sativa (рис) |
480 000 000 |
57 000 |
|
|
Mus musculus (мышь) |
2 634 266 500 |
27 000 |
|
|
Homo sapiens (человек) |
3 070 128 600 |
29 000 |
Примечание. Информация постоянно обновляется; для получения более свежей информации обратитесь к сайтам, посвященным отдельным геномным проектам
* Для всех эукариот, кроме дрожжей, приводится диплоидный набор хромосом. Диплоидный
набор хромосом (от греч. diploos- двойной и eidos- вид) - двойной набор хромосом (2n), каждая из которых имеет себе гомологичную.
**Гаплоидный набор. Дикие штаммы дрожжей обычно имеют восемь (октаплоидный) или больше наборов таких хромосом.
***Для самок с двумя Х хромосомами. У самцов есть Х хромосома, но нет Y, т. е. всего 11 хромосом.
В клетке дрожжей, одних из самых маленьких эукариот, в 2,6 раза больше ДНК, чем в клетке E. coli (табл. 2). Клетки плодовой мушки Drosophila , классического объекта генетических исследований, содержат в 35 раз больше ДНК, а клетки человека — примерно в 700 раз больше ДНК, чем клетки E. coli. Многие растения и амфибии содержат еще больше ДНК. Генетический материал клеток эукариот организован в виде хромосом. Диплоидный набор хромосом (2n ) зависит от вида организма (табл. 2).
Например, в соматической клетке человека 46 хромосом (рис. 17 ). Каждая хромосома эукариотической клетки, как показано на рис. 17, а , содержит одну очень крупную двухспиральную молекулу ДНК. Двадцать четыре хромосомы человека (22 парные хромосомы и две половые хромосомы X и Y) различаются по длине более чем в 25 раз. Каждая хромосома эукариот содержит определенный набор генов.

Рис. 17. Хромосомы эукариот. а — пара связанных и конденсированных сестринских хроматид из хромосомы человека. В такой форме эукариотические хромосомы пребывают после репликации и в метафазе в процессе митоза. б — полный набор хромосом из лейкоцита одного из авторов книги. В каждой нормальной соматической клетке человека содержится 46 хромосом.
Если соединить между собой молекулы ДНК человеческого генома (22 хромосомы и хромосомы X и Y или Х и Х), получится последовательность длиной около одного метра. Прим.: У всех млекопитающих и других организмов с гетерогаметным мужским полом, у самок две X-хромосомы (XX), а у самцов — одна X-хромосома и одна Y-хромосома (XY).
Большинство клеток человека , поэтому общая длина ДНК таких клеток около 2м. У взрослого человека примерно 10 14 клеток, таким образом, общая длина всех молекул ДНК составляет 2・10 11 км. Для сравнения, окружность Земли — 4・10 4 км, а расстояние от Земли до Солнца — 1,5・10 8 км. Вот как удивительно компактно упакована ДНК в наших клетках!
В клетках эукариот есть и другие органеллы, содержащие ДНК, — это митохондрии и хлоропласты. Выдвигалось множество гипотез относительно происхождения ДНК митохондрий и хлоропластов. Общепризнанная сегодня точка зрения заключается в том, что они представляют собой рудименты хромосом древних бактерий, которые проникли в цитоплазму хозяйских клеток и стали предшественниками этих органелл. Митохондриальная ДНК кодирует митохондриальные тРНК и рРНК, а также несколько митохондриальных белков. Более 95% митохондриальных белков кодируется ядерной ДНК.
СТРОЕНИЕ ГЕНОВ
Рассмотрим строение гена у прокариот и эукариот, их сходства и различия. Несмотря на то, что ген — это участок ДНК, кодирующий всего один белок или РНК, кроме непосредственно кодирующей части, он также включает в себя регуляторные и иные структурные элементы, имеющие разное строение у прокариот и эукариот.
Кодирующая последовательность - основная структурно-функциональная единица гена, именно в ней находятся триплеты нуклеотидов, кодирующие аминокислотную последовательность. Она начинается со старт-кодона и заканчивается стоп-кодоном.
До и после кодирующей последовательности находятся нетранслируемые 5’- и 3’-последовательности . Они выполняют регуляторные и вспомогательные функции, например, обеспечивают посадку рибосомы на и-РНК.
Нетранслируемые и кодирующая последовательности составлют единицу транскрипции - транскрибируемый участок ДНК, то есть участок ДНК, с которого происходит синтез и-РНК.
Терминатор - нетранскрибируемый участок ДНК в конце гена, на котором останавливается синтез РНК.
В начале гена находится регуляторная область , включающая в себя промотор и оператор .
Промотор - последовательность, с которой связывается полимераза в процессе инициации транскрипции. Оператор - это область, с которой могут связываться специальные белки - репрессоры , которые могут уменьшать активность синтеза РНК с этого гена - иначе говоря, уменьшать его экспрессию .
Строение генов у прокариот
Общий план строения генов у прокариот и эукариот не отличается - и те, и другие содержат регуляторную область с промотором и оператором, единицу транскрипции с кодирующей и нетранслируемыми последовательностями и терминатор. Однако организация генов у прокариот и эукариот отличается.

Рис. 18. Схема строения гена у прокариот (бактерий) - изображение увеличивается
В начале и в конце оперона есть единые регуляторные области для нескольких структурных генов. С транскрибируемого участка оперона считывается одна молекула и-РНК, которая содержит несколько кодирующих последовательностей, в каждой из которых есть свой старт- и стоп-кодон. С каждого из таких участков с интезируется один белок. Таким образом, с одной молекулы и-РНК синтезируется несколько молекул белка.
Для прокариот характерно объединение нескольких генов в единую функциональную единицу - оперон . Работу оперона могут регулировать другие гены, которые могут быть заметно удалены от самого оперона - регуляторы . Белок, транслируемый с этого гена называется репрессор . Он связывается с оператором оперона, регулируя экспрессию сразу всех генов, в нем содержащихся.
Для прокариот также характерно явление сопряжения транскрипции и трансляции .
![]()
Рис. 19 Явление сопряжения транскрипции и трансляции у прокариот - изображение увеличивается
Такое сопряжение не встречается у эукариот из-за наличия у них ядерной оболочки, отделяющей цитоплазму, где происходит трансляция, от генетического материала, на котором происходит транскрипция. У прокариот во время синтеза РНК на матрице ДНК с синтезируемой молекулой РНК может сразу связываться рибосома. Таким образом, трансляция начинается еще до завершения транскрипции. Более того, с одной молекулой РНК может одновременно связываться несколько рибосом, синтезируя сразу несколько молекул одного белка.
Строение генов у эукариот
Гены и хромосомы эукариот очень сложно организованы
У бактерий многих видов всего одна хромосома, и почти во всех случаях в каждой хромосоме присутствует по одной копии каждого гена. Лишь немногие гены, например гены рРНК, содержатся в нескольких копиях. Гены и регуляторные последовательности составляют практически весь геном прокариот. Более того, почти каждый ген строго соответствует аминокислотной последовательности (или последовательности РНК), которую он кодирует (рис. 14).
Структурная и функциональная организация генов эукариот гораздо сложнее. Исследование хромосом эукариот, а позднее секвенирование полных последовательностей геномов эукариот принесло много сюрпризов. Многие, если не большинство, генов эукариот обладают интересной особенностью: их нуклеотидные последовательности содержат один или несколько участков ДНК, в которых не кодируется аминокислотная последовательность полипептидного продукта. Такие нетранслируемые вставки нарушают прямое соответствие между нуклеотидной последовательностью гена и аминокислотной последовательностью кодируемого полипептида. Эти нетранслируемые сегменты в составе генов называют интронами , или встроенными последовательностями , а кодирующие сегменты — экзонами . У прокариот лишь немногие гены содержат интроны.
Итак, у эукариот практически не встречается объединение генов в опероны, и кодирующая последовательность гена эукариот чаще всего разделена на транслируемые участки - экзоны , и нетранслируемые участки - интроны.
В большинстве случаев функция интронов не установлена. В целом, лишь около 1,5% ДНК человека являются ≪кодирующими≫, т. е. несут информацию о белках или РНК. Однако с учетом крупных интронов получается, что ДНК человека на 30% состоит из генов. Поскольку гены составляют относительно небольшую долю в геноме человека, значительная часть ДНК остается неучтенной.

Рис. 16. Схема строение гена у эукариот - изображение увеличивается
С каждого гена сначала синтезируется незрелая, или пре-РНК, которая содержит в себе как интроны, так и экзоны.
После этого проходит процесс сплайсинга, в результате которого интронные участки вырезаются, и образуется зрелая иРНК, с которой может быть синтезирован белок.

Рис. 20. Процесс альтернативного сплайсинга - изображение увеличивается
Такая организация генов позволяет, например, осуществить , когда с одного гена могут быть синтезированы разные формы белка, за счет того, что в процессе сплайсинга экзоны могут сшиваться в разных последовательностях.

Рис. 21. Отличия в строении генов прокариот и эукариот - изображение увеличивается
МУТАЦИИ И МУТАГЕНЕЗ
Мутацией называется стойкое изменение генотипа, то есть изменение нуклеотидной последовательности.
Процесс, который приводит к возникновению мутаций называется мутагенезом , а организм, все клетки которого несут одну и ту же мутацию — мутантом .
Мутационная теория была впервые сформулирована Гуго де Фризом в 1903 году. Современный ее вариант включает в себя следующие положения:
1. Мутации возникают внезапно, скачкообразно.
2. Мутации передаются из поколения в поколение.
3. Мутации могут быть полезными, вредными или нейтральными, доминантными или рецессивными.
4. Вероятность обнаружения мутаций зависит от числа исследованных особей.
5. Сходные мутации могут возникать повторно.
6. Мутации не направленны.
Мутации могут возникать под действием различных факторов. Различают мутации, возникшие под действием мутагенных воздействий : физических (например, ультрафиолета или радиации), химических (например, колхицина или активных форм кислорода) и биологических (например, вирусов). Также мутации могут быть вызваны ошибками репликации .
В зависимости от условий появления мутации подразделяют на спонтанные — то есть мутации, возникшие в нормальных условиях, и индуцированые — то есть мутации, которые возникли при особых условиях.
Мутации могут возникать не только в ядерной ДНК, но и, например, в ДНК митохондрий или пластид. Соответственно, мы можем выделять ядерные и цитоплазматические мутации.
В результате возникновения мутаций часто могут появляться новые аллели. Если мутантный аллель подавляет действие нормального, мутация называется доминантной . Если нормальный аллель подавляет мутантный, такая мутация называется рецессивной . Большинство мутаций, приводящих к возникновению новых аллелей являются рецессивными.
По эффекту выделяют мутации адаптивные , приводящие к повышению приспособленности организма к среде, нейтральные , не влияющие на выживаемость, вредные , понижающие приспособленность организмов к условиям среды и летальные , приводящие к смерти организма на ранних стадиях развития.
По последствиям выделяются мутации, приводящие к потери функции белка , мутации, приводящие к возникновению у белка новой функции , а также мутации, которые изменяют дозу гена , и, соответственно, дозу белка синтезируемого с него.
Мутация может возникнуть к любой клетке организма. Если мутация возникает в половой клетке, она называется герминативной (герминальной, или генеративной). Такие мутации не проявляются у того организма, у которого они появились, но приводят к появлению мутантов в потомстве и передаются по наследству, поэтому они важны для генетики и эволюции. Если мутация возникает в любой другой клетке, она называется соматической . Такая мутация может в той или иной степени проявляться у того организма, у которого она возникла, например, приводить к образованию раковых опухолей. Однако такая мутация не передается по наследству и не влияет на потомков.
Мутации могут затрагивать разные по размеру участки генома. Выделяют генные , хромосомные и геномные мутации.
Генные мутации
Мутации, которые возникают в масштабе меньшем, чем один ген, называются генными , или точечными (точковыми) . Такие мутации приводят к изменению одного и нескольких нуклеотидов в последовательности. Среди генных мутаций выделяют замены , приводящие к замене одного нуклеотида на другой, делеции , приводящие к выпадению одного из нуклеотидов, инсерции , приводящие к добавлению лишнего нуклеотида в последовательность.

Рис. 23. Генные (точечные) мутации
По механизму воздействия на белок, генные мутации делят на: синонимичные , которые (в результате вырожденности генетического кода) не приводят к изменению аминокислотного состава белкового продукта, миссенс-мутации , которые приводят к замене одной аминокислоты на другую и могут влиять на структуру синтезируемого белка, хотя часто они оказываются незначительными, нонсенс-мутации , приводящие к замене кодирующего кодона на стоп-кодон, мутации, приводящие к нарушению сплайсинга:

Рис. 24. Схемы мутаций
Также по механизму воздействия на белок выделяют мутации, приводящие к сдвигу рамки
считывания
, например, инсерции и делеции. Такие мутации, как и нонсенс-мутации, хоть и возникают в одной точке гена, часто воздействуют на всю структуру белка, что может привести к полному изменению его структуры.

Рис. 29. Хромосома до и после дупликации
Геномные мутации
Наконец, геномные мутации затрагивают весь геном целиком, то есть меняется количество хромосом. Выделяют полиплоидии — увеличение плоидности клетки, и анеуплоидии, то есть изменение количества хромосом, например, трисомии (наличие у одной из хромосом дополнительного гомолога) и моносомии (отсутствие у хромосомы гомолога).
Видео по теме ДНК
РЕПЛИКАЦИЯ ДНК, КОДИРОВАНИЕ РНК, СИНТЕЗ БЕЛКА
Нуклеиновые кислоты - высокомолекулярные вещества, состоящие из мононуклеотидов, которые соединены друг с другом в полимерную цепочку с помощью 3",5"- фосфодиэфирных связей и упакованы в клетках определенным образом.
Нуклеиновые кислоты - биополимеры двух разновидностей: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). Каждый биополимер состоит из нуклеотидов, различающихся по углеводному остатку (рибозе, дезоксирибозе) и одному из азотистых оснований (урацил, тимин). Соответственно этим различиям нуклеиновые кислоты и получили свое название.
Структура дезоксирибонуклеиновой кислоты
Нуклеиновые кислоты имеют первичную, вторичную и третичную структуру.
Первичная структура ДНК
Первичной структурой ДНК называют линейную полинуклеотидную цепь, в которой мононуклеотиды соединены 3", 5"-фосфодиэфирными связями. Исходным материалом при сборке цепи нуклеиновой кислоты в клетке является нуклеозид 5"-трифосфат, который в результате удаления β и γ остатков фосфорной кислоты способен присоединить 3"-атом углерода другого нуклеозида. Таким образом, 3"-атом углерода одной дезоксирибозы ковалентно связывается с 5"-атомом углерода другой дезоксирибозы посредством одного остатка фосфорной кислоты и образует линейную полинуклеотидную цепь нуклеиновой кислоты. Отсюда и название: 3", 5"-фосфодиэфирные связи. Азотистые основания не принимают участия в соединении нуклеотидов одной цепи (рис. 1.).
Такое соединение, между остатком молекулы фосфорной кислоты одного нуклеотида и углеводом другого, приводит к образованию пентозо-фосфатного скелета молекулы полинуклеотида, на котором сбоку один за другим присоединяются азотистые основания. Их последовательность расположения в цепях молекул нуклеиновых кислот строго специфична для клеток разных организмов, т.е. носит видовой характер (правило Чаргаффа).
Линейная цепь ДНК, длина которой зависит от числа входящих в цепь нуклеотидов, имеет два конца: один называется 3"-концом и содержит свободный гидроксил, а другой - 5"-концом, содержит остаток фосфорной кислоты. Цепь полярна и может иметь напрвление 5"->3" и 3"->5". Исключением являются кольцевые ДНК.
Генетический "текст" ДНК составлен с помощью кодовых "слов" - триплетов нуклеотидов, называемых кодонами. Участки ДНК, содержащие информацию о первичной структуре всех типов РНК, называют структурными генами.
Полинуклеодитные цепочки ДНК достигают гигантских размеров, поэтому в клетке они упакованы определенным образом.
Изучая состав ДНК, Чаргафф (1949) установил важные закономерности, касающиеся содержания отдельных оснований ДНК. Они помогли раскрыть вторичную структуру ДНК. Эти закономерности называют правилами Чаргаффа. Правила Чаргаффа
Эти правила говорят о том, что при построении ДНК должно соблюдаться довольно строгое соответствие (спаривание) не пуриновых и пиримидиновых оснований вообще, а конкретно тимина с аденином и цитозина с гуанином. На основании этих правил в том числе, в 1953 г. Уотсон и Крик предложили модель вторичной структуры ДНК, получившую название двойной спирали (рис.). |

Вторичная структура ДНК
Вторичная структура ДНК - это двойная спираль, модель которой была предложена Д.Уотсоном и Ф.Криком в 1953 году.
Предпосылки к созданию модели ДНК
В результате первоначальных анализов сложилось представление, что ДНК любого происхождения содержит все четыре нуклеотида в равных молярных количествах. Однако в 1940-х годах Э. Чаргафф и его сотрудники в результате анализа ДНК, выделенных из разнообразных организмов, ясно показали, что азотистые основания содержатся в них в различных количественных соотношениях. Чаргафф нашел, что, хотя эти соотношения одинаковы для ДНК из всех клеток одного и того же вида организмов, ДНК от разных видов могут заметно различаться по содержанию тех или иных нуклеотидов. Это наводило на мысль, что различия в соотношении азотистых оснований, возможно, связаны с каким-то биологическим кодом. Хотя соотношение отдельных пуриновых и пиримидиновых оснований в различных образцах ДНК оказалось неодинаковым, при сравнении результатов анализов выявилась определенная закономерность: во всех образцах общее количество пуринов было равно общему количеству пиримидинов (А + Г = Т + Ц), количество аденина - количеству тимина (А = Т), а количество гуанина - количеству цитозина (Г = Ц). ДНК, выделенная из клеток млекопитающих, была в целом богаче аденином и тимином и относительно беднее гуанином и цитозином, тогда как у бактерий ДНК была богаче гуанином и цитозином и относительно беднее аденином и тимином. Эти данные составили важную часть фактического материала, на основе которого позднее была построена модель структуры ДНК Уотсона - Крика.
Еще одним важным косвенным указанием на возможную структуру ДНК послужили данные Л. Полинга о строении белковых молекул. Полинг показал, что возможно несколько различных устойчивых конфигураций аминокислотной цепи в белковой молекуле. Одна из распространенных конфигураций пептидной цепи - α-спираль - представляет собой правильную винтообразную структуру. При такой структуре возможно образование водородных связей между аминокислотами, находящимися на смежных витках цепи. Полинг описал α-спиральную конфигурацию полипептидной цепи в 1950 году и высказал предположение, что и молекулы ДНК, вероятно, имеют спиральную структуру, закрепленную водородными связями.
Однако наиболее ценные сведения о строении молекулы ДНК дали результаты рентгеноструктурного анализа. Рентгеновские лучи, проходя сквозь кристалл ДНК, претерпевают дифракцию, т. е. отклоняются в определенных направлениях. Степень и характер отклонения лучей зависят от структуры самих молекул. Дифракционная рентгенограмма (рис. 3) дает опытному глазу ряд косвенных указаний относительно строения молекул исследуемого вещества. Анализ дифракционных рентгенограмм ДНК привел к заключению, что азотистые основания (имеющие плоскую форму) уложены наподобие стопки тарелок. Рентгенограммы позволили выявить в структуре кристаллической ДНК три главных периода: 0,34, 2 и 3,4 нм.

Модель ДНК Уотсона-Крика
Исходя из аналитических данных Чаргаффа, рентгенограмм, полученных Уилкинсом и исследований химиков, предоставивших сведения о точных расстояниях между атомами в молекуле, об углах между связями данного атома и о величине атомов, Уотсон и Крик начали строить физические модели отдельных составных частей молекулы ДНК в определенном масштабе и "подгонять" их друг к другу с таким расчетом, чтобы полученная система соответствовала различным экспериментальным данным [показать] .
Еще раньше было известно, что в цепи ДНК соседние нуклеотиды соединены фосфодиэфирными мостиками, связывающими 5"-углеродный атом дезоксирибозы одного нуклеотида с 3"-углеродным атомом дезоксирибозы следующего нуклеотида. Уотсон и Крик не сомневались в том, что период 0,34 нм соответствует расстоянию между последовательными нуклеотидами в цепи ДНК. Далее, можно было предполагать, что период 2 нм соответствует толщине цепи. А для того чтобы объяснить, какой реальной структуре соответствует период 3,4 нм, Уотсон и Крик, так же как ранее Полинг, предположили, что цепь закручена в виде спирали (или, точнее, образует винтовую линию, так как спираль в строгом смысле этого слова получается тогда, когда витки образуют в пространстве коническую, а не цилиндрическую поверхность). Тогда период 3,4 нм будет соответствовать расстоянию между последовательными витками этой спирали. Такая спираль может быть очень плотной или же несколько растянутой, т. е. витки ее могут быть пологими или крутыми. Поскольку период 3,4 нм ровно в 10 раз больше расстояния между последовательными нуклеотидами (0,34 нм), ясно, что каждый полный виток спирали содержит 10 нуклеотидов. По этим данным Уотсон и Крик смогли вычислить плотность полинуклеотидной цепи, закрученной в спираль диаметром 2 нм, с расстоянием между витками, равным 3,4 нм. Оказалось, что у такой цепи плотность была бы вдвое меньше фактической плотности ДНК, которая была уже известна. Пришлось предположить, что молекула ДНК состоит из двух цепей - что это двойная спираль из нуклеотидов.
Следующей задачей было, конечно, выяснение пространственных отношений между обеими цепями, образующими двойную спираль. Испробовав на своей физической модели ряд вариантов расположения цепей, Уотсон и Крик нашли, что всем имеющимся данным лучше всего соответствует такой вариант, в котором две полинуклеотидные спирали идут в противоположных направлениях; при этом цепи, состоящие из остатков сахара и фосфата, образуют поверхность двойной спирали, а пурины и пиримидины располагаются внутри. Расположенные друг против друга основания, принадлежащие двум цепям, попарно соединены водородными связями; именно эти водородные связи и удерживают цепи вместе, фиксируя таким образом общую конфигурацию молекулы.
Двойную спираль ДНК можно представить себе в виде винтообразно закрученной веревочной лестницы, так чтобы перекладины ее оставались в горизонтальном положении. Тогда две продольные веревки будут соответствовать цепям из остатков сахара и фосфата, а перекладины - парам азотистых оснований, соединенных водородными связями.
В результате дальнейшего изучения возможных моделей Уотсон и Крик пришли к выводу, что каждая "перекладина" должна состоять из одного пурина и одного пиримидина; при периоде 2 нм (что соответствует диаметру двойной спирали) для двух пуринов не хватило бы места, а два пиримидина не могли бы при этом располагаться достаточно близко друг к другу, чтобы образовать надлежащие водородные связи. Углубленное исследование детальной модели показало, что аденин и цитозин, составляя подходящую по размерам комбинацию, все же не могли бы располагаться таким образом, чтобы между ними образовались водородные связи. Аналогичные сообщения заставили исключить также комбинацию гуанин - тимин, тогда как сочетания аденин - тимин и гуанин - цитозин оказались вполне приемлемыми. Природа водородных связей такова, что аденин образует пару с тимином, а гуанин - с цитозином. Это представление о специфическом спаривании оснований позволяло объяснить "правило Чаргаффа", согласно которому в любой молекуле ДНК количество аденина всегда равно содержанию тимина, а количество гуанина - количеству цитозина. Между аденином и тимином образуются две водородные связи, а между гуанином и цитозином - три. Благодаря этой специфичности в образовании водородных связей против каждого аденина в одной цепи в другой оказывается тимин; точно так же против каждого гуанина может находиться только цитозин. Таким образом, цепи комплементарны друг другу, т. е. последовательность нуклеотидов в одной цепи однозначно определяет их последовательность в другой. Две цепи идут в противоположных направлениях, и их концевые фосфатные группы находятся на противоположных концах двойной спирали.
В результате своих исследований, в 1953 году Уотсон и Крик предложили модель строения молекулы ДНК (рис. 3), которая остается актуальной по настоящее время. Согласно модели молекула ДНК состоит из двух комплементарных полинуклеотидных цепей. Каждая цепь ДНК представляет полинуклеотид, состоящий из нескольких десятков тысяч нуклеотидов. В ней соседние нуклеотиды образуют регулярный пентозо-фосфатный остов за счет соединения остатка фосфорной кислоты и дезоксирибозы прочной ковалентной связью. Азотистые основания одной полинуклеотидной цепи при этом располагаются в строго определенном порядке против азотистых оснований другой. Чередование азотистых оснований в полинуклеотидной цепи нерегулярно.
Расположение азотистых оснований в цепи ДНК является комплементарным (от греч. "комплемент" - дополнение), т.е. против аденина (А) всегда оказывается тимин (Т), а против гуанина (Г) - только цитозин (Ц). Это объясняется тем, что А и Т, а также Г и Ц строго соответствуют друг другу, т.е. дополняют друг другу. Такое соответствие задается химической структурой оснований, позволяющей образовать водородные связи в паре пурина и пиримидина. Между А и Т возникают две связи, между Г и Ц - три. Эти связи обеспечивают частичную стабилизацию молекулы ДНК в пространстве. Устойчивость двойной спирали при этом прямо пропорциональна числу связей G≡С, являющихся более стабильными по сравнению со связями А=Т.
Известная последовательность расположения нуклеотидов в одной цепи ДНК позволяет по принципу комплементарности установить нуклеотиды другой цепи.
Кроме того, установлено, что азотистые основания, имеющие ароматическую структуру, в водном растворе располагаются один над другим, формируя как бы стопку монет. Такой процесс формирования стопок из органических молекул называется стекинг. Полинуклеотидные цепи молекулы ДНК рассматриваемой модели Уотсона-Крика имеют аналогичное физико-химическое состояние, их азотистые основания располагаются в виде стопки монет, между плоскостями которых возникают ван-дер-ваальсовы взаимодействия (стекинг-взаимодействия).
Водородные связи между комплементарными основаниями (по горизонтали) и стекинг-взаимодействие между плоскостями оснований в полинуклеотидной цепи за счет ван-дер-ваальсовых сил (по вертикали) обеспечивает молекуле ДНК дополнительную стабилизацию в пространстве.
Сахарофосфатные остовы обеих цепей обращены наружу, а основания внутрь, навстречу друг другу. Направление цепей в ДНК антипараллельно (одна из них имеет направление 5"->3", другая - 3"->5", т.е. 3"-конец одной цепи расположен напротив 5"-конца другой.). Цепи образуют правые спирали с общей осью. Один виток спирали составляет 10 нуклеотидов, размер витка 3,4 нм, высота каждого нуклеотида 0,34 нм, диаметр спирали – 2,0 нм. В результате вращения одной цепи вокруг другой, образуется большая борозда (диаметром около 20 Å) и малая борозда (около 12 Å) двойной спирали ДНК. Такая форма двойной спирали Уотсона-Крика в дальнейшем получила название В-формы. В клетках ДНК обычно существует в В-форме, которая является самой стабильной.
Функции ДНК
Предложенная модель объясняла многие биологические свойства дезоксирибонуклеиновой кислоты, в том числе хранение генетической информации и многообразие генов, обеспечиваемое большим разнообразием последовательных сочетаний 4-х нуклеотидов и фактом существования генетического кода, способность к самовоспроизведению и передаче генетической информации, обеспечиваемое процессом репликации, и реализацию генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов.
Oсновные функции ДНК.
- ДНК является носителем генетической информации, что обеспечивается фактом существования генетического кода.
- Воспроизведение и передана генетической информации в поколениях клеток и организмов. Эта функция обеспечивается процессом репликации.
- Реализация генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов. Эта функция обеспечивается процессами транскрипции и трансляции.

Формы организации двухцепочечной ДНК
ДНК может формировать несколько типов двойных спиралей (рис.4). В настоящее время уже известно шесть форм (от А до Е и Z-форма).
Структурные формы ДНК, как установила Розалинда Франклин, зависят от насыщения водой молекулы нуклеиновой кислоты. В исследованиях волокон ДНК при помощи рентгеноструктурного анализа было показано, что рентгенограмма радикальным образом зависит от того, при какой относительной влажности, при какой степени насыщения водой этого волокна происходит эксперимент. Если волокно было достаточно насыщено водой, то получалась одна рентгенограмма. При высушивании возникала совершенно другая рентгенограмма, сильно отличающаяся от рентгенограммы волокна высокой влажности.
Молекула ДНК высокой влажности получила название В-формы . При физиологических условиях (низкая концентрация соли, высокая степерь гидратации) доминирующим структурным типом ДНК является В-форма (основная форма двухцепочечной ДНК - модель Уотсона-Крика). Шаг спирали такой молекулы равен 3,4 нм. На виток приходится 10 комплементарных пар в виде скрученных стопок "монет" - азотистых оснований. Стопки удерживаются водородными связями между двумя противолежащими "монетами" стопок, и "обмотаны" двумя лентами фосфодиэфирного остова, закрученными в правую спираль. Плоскости азотистых оснований перпендикулярны оси спирали. Соседние комплементарные пары повернуты друг относительно друга на 36°. Диаметр спирали 20Å, причем пуриновый нуклеотид занимает 12Å, а пиримидиновый - 8Å.
Молекула ДНК более низкой влажности получила название А-формы . А-форма образуется в условиях менее высокой гидратации и при более высоком содержании ионов Na + или К + . Эта более широкая правоспиральная конформация имеет 11 пар азотистых оснований на виток. Плоскости азотистых оснований имеют более сильный наклон к оси спирали, они отклонены от нормали к оси спирали на 20°. Отсюда следует наличие внутренней пустоты диаметром 5Å. Расстояние между соседними нуклеотидами составляет 0,23 нм, длина витка – 2,5 нм, диаметр спирали – 2,3 нм.
Первоначально считали, что А-форма ДНК менее важна. Однако в дальнейшем выяснилось, что А-форма ДНК, также как и В-форма, имеет огромное биологическое значение. А-форму имеет спираль РНК-ДНК в комплексе матрица-затравка, а также спираль РНК-РНК и шпилечные структуры РНК (2’-гидроксильная группа рибозы не позволяет молекулам РНК образовывать В-форму). А-форма ДНК обнаружена в спорах. Установлено, что А-форма ДНК в 10 раз устойчивее к действию УФ-лучей, чем В-форма.
А-форму и В-форму называют каноническими формами ДНК.
Формы С-Е также правоспиральные, их образование можно наблюдать только в специальных экспериментах, и, по-видимому, они не существуют in vivo. С-форма ДНК имеет структуру, сходную с В-ДНК. Число пар оснований на виток составляет 9,33, длина витка спирали равна 3,1 нм. Пары оснований наклонены на угол 8 градусов относительно перпендикулярного положения к оси. Желобки по размерам близки к желобкам В-ДНК. При этом главный желобок несколько мельче, а минорный желобок – глубже. В С-форму могут переходить природные и синтетические полинуклеотиды ДНК.
| Таблица 1. Характеристика некоторых типов структур ДНК | |||
| Тип спирали | A | B | Z |
| Шаг спирали | 0,32 нм | 3,38 нм | 4,46 нм |
| Закрученность спирали | Правая | Правая | Левая |
| Число пар оснований на виток | 11 | 10 | 12 |
| Расстояние между плоскостями оснований | 0,256 нм | 0,338 нм | 0,371 нм |
| Конформация гликозидной связи | анти | анти | анти-С син-Г |
| Конформация фуранозного цикла | С3"-эндо | С2"-эндо | С3"-эндо-Г С2"-эндо-Ц |
| Ширина бороздки, малой/большой | 1,11/0,22 нм | 0,57/1,17 нм | 0,2/0,88 нм |
| Глубина бороздки, малой/большой | 0,26/1,30 нм | 0,82/0,85 нм | 1,38/0,37 нм |
| Диаметр спирали | 2,3 нм | 2,0 нм | 1,8 нм |
Структурные элементы ДНК
(неканонические структуры ДНК)
К структурным элементам ДНК можно отнести необычные структуры, ограниченные какими-то специальными последовательностями:
|

Z-форма ДНК была открыта в 1979 году при изучении гексануклеотида d(CG)3 - . Ее открыл профессор Массачусетского технологического института Александр Рич с сотрудниками. Z-форма стала одним из важнейших структурных элементов ДНК в связи с тем, что ее образование наблюдалось в участках ДНК, где пурины чередуются с пиримидинами (например, 5’-ГЦГЦГЦ-3’), или в повторах 5’-ЦГЦГЦГ-3’, содержащих метилированный цитозин. Существенным условием образования и стабилизации Z-ДНК являлось присутствие в ней пуриновых нуклеотидов в син-конформации, чередующихся с пиримидиновыми основаниями в анти-конформации.
Природные молекулы ДНК в основном существуют в правой В-форме, если они не содержат последовательностей типа (ЦГ)n. Однако, если такие последовательности входят в состав ДНК, то эти участки при изменении ионной силы раствора или катионов, нейтрализующих отрицательный заряд на фосфодиэфирном каркасе, могут переходить в Z-форму, при этом другие участки ДНК в цепи остаются в классической В-форме. Возможность такого перехода указывает на то, что две цепи в двойной спирали ДНК находятся в динамическом состоянии и могут раскручиваться друг относительно друга, переходя из правой формы в левую и наоборот. Биологические следствия такой лабильности, допускающей конформационные превращения структуры ДНК пока не вполне понятны. Полагают, что участки Z-ДНК играют определенную роль в регуляции экспрессии некоторых генов и принимают участие в генетической рекомбинации.
Z-форма ДНК - это левозакрученная двойная спираль, в которой фосфодиэфирный остов расположен зигзагообразно вдоль оси молекулы. Отсюда и название молекулы (zigzag)-ДHK. Z-ДНК - наименее скрученная (12 пар оснований на виток) и наиболее тонкая из известных в природе. Расстояние между соседними нуклеотидами составляет 0,38 нм, длина витка – 4,56 нм, диаметр Z-ДНК – 1,8 нм. Кроме того, внешний вид этой молекулы ДНК отличается наличием одной бороздки.
Z-форма ДНК была обнаружена в клетках прокариот и эукариот. В настоящее время получены антитела, способные отличать Z-форму от В-формы ДНК. Эти антитела связываются с определенными участками гигантских хромосом клеток слюнных желез дрозофилы (Dr. melanogaster). За реакцией связывания легко следить из-за необычного строения этих хромосом, у которых более плотные участки (диски) контрастируют с менее плотными (междисками). Участки Z-ДНК расположены в междисках. Из этого следует, что Z-форма реально существует в естественных условиях, хотя размеры индивидуальных участков Z-формы пока неизвестны.

(перевертыши) - наиболее известные и часто встречающиеся в ДНК последовательности оснований. Палиндромом называют слово или фразу, которое читается слева направо и наоборот одинаково. Примерами таких слов или фраз являются: ШАЛАШ, КАЗАК, ПОТОП, А РОЗА УПАЛА НА ЛАПУ АЗОРА. В применении к участкам ДНК данный термин (палиндром) означает одинаковое чередование нуклеотидов вдоль цепи справа налево и слева направо (подобно буквам в слове "шалаш" и пр.).
Палиндром характеризуется наличием инвертированных повторов последовательностей оснований имеющих симметрию второго порядка относительно двух цепей ДНК. Такие последовательности, по вполне понятной причине, являются самокомплементарными и имеют склонность к образованию шпилечных или крестообразных структур (рис.). Шпильки помогают регуляторным белкам узнавать место списывания генетического текста ДНК хромосом.
В тех случаях, когда инвертированный повтор присутствует в одной и той же цепи ДНК такая последовательность называется зеркальным повтором. Зеркальные повторы не обладают свойствами самокомплементарности и, поэтому не способны к формированию шпилечных или крестообразных структур. Последовательности такого типа обнаружены практически во всех крупных молекулах ДНК и могут включать от всего нескольких пар оснований до нескольких тысяч пар оснований.
Присутствие палиндромов в виде крестообразных структур в эукариотических клетках не доказано, хотя некоторое количество крестообразных структур обнаружено в условиях in vivo в клетках E. coli. Наличие в составе РНК или одноцепочечных ДНК самокомплементарных последовательностей служит основной причиной сворачивания в растворах нуклеиновой цепи в определенную пространственную структуру, отличающуюся формированием множества "шпилек".

Н-форма ДНК - это спираль, которую образуют три цепи ДНК - тройная спираль ДНК. Представляет собой комплекс уотсон-криковской двойной спирали с третьей одноцепочечной нитью ДНК, которая укладывается в ее большой желобок, с образованием так называемой хугстиновской пары.
Образование подобного триплекса происходит в результате сложения двойной спирали ДНК таким образом, что половина ее участка остается в виде двойной спирали, а вторая половина разъединяется. При этом одна из разъединенных спиралей образует новую структуру с первой половиной двойной спирали - тройную спираль, а вторая оказывается неструктурированной, в виде однонитевого участка. Особенностью этого структурного перехода является резкая зависимость от рН среды, протоны которой стабилизируют новую структуру. В силу этой особенности новая структура получила название Н-формы ДНК, образование которой было обнаружено в сверхспирализованных плазмидах, содержащих гомопурин-гомопиримидиновые участки, представляющие собой зеркальный повтор.
В дальнейших исследованиях была установлена возможность осуществления структурного перехода некоторых гомопурин-гомопиримидиновых двунитиевых полинуклеотидов с образованием трехнитиевой структуры, содержащей:
- одну гомопуриновую и две гомопиримидиновые нити (Py-Pu-Py триплекс
) [хугстиновское взаимодействие].
Составляющие блоки Py-Pu-Py триплекса - канонические изоморфные CGC+ и TAT триады. Стабилизация триплекса требует протонирования триады CGC+, поэтому эти триплексы зависят от рН раствора.
- одну гомопиримидиновую и две гомопуриновые нити (Py-Pu-Pu триплекс
) [обратное хугстиновское взаимодействие].
Составляющие блоки Py-Pu-Pu триплекса - канонические изоморфные CGG и TAA триад. Существенным свойством Py-Pu-Pu триплексов является зависимость их стабильности от присутствия двухзарядных ионов, причем для стабилизации триплексов разной последовательности необходимы различные ионы. Поскольку для образования Py-Pu-Pu триплексов не требуется протонирования входящих в их состав нуклеотидов, такие триплексы могут существовать при нейтральных pH.
Прим.: прямое и обратное хугстиновское взаимодействие объясняется симметрией 1-метилтимина: поворот на 180° приводит к тому, что место атома О4 занимает атом О2, при этом система водородных связей сохраняется.
Известны два вида тройных спиралей:
- параллельные тройные спирали, в которых полярность третьей цепи совпадает с полярностью гомопуриновой цепи Уотсон-криковского дуплекса
- антипараллельные тройные спирали, в которых полярности третьей и гомопуриновой цепей противоположны.

G-квадруплекс - 4-х спиральная ДНК. Такая структура образуется в случае, если имеются четыри гуанина, которые образуют так называемый G-квадруплекс - хоровод из четырех гуанинов.
Первые намеки на возможность образования таких структур были получены задолго до прорывной работы Уотсона и Крика - еще в 1910 году. Тогда немецкий химик Ивар Банг обнаружил, что один из компонентов ДНК - гуанозиновая кислота - при высоких концентрациях образует гели, в то время как другие составные части ДНК таким свойством не обладают.
В 1962 году с помощью рентгеноструктурного метода удалось установить структуру ячейки этого геля. Она оказалась составлена из четырех остатков гуанина, связывающих друг друга по кругу и образующих характерный квадрат. В центре связь поддерживает ион металла (Na, K, Mg). Такие же структуры могут образовываться и в ДНК, если в ней много гуанина. Эти плоские квадраты (G-квартеты) складываются в стопки, и получаются довольно устойчивые, плотные структуры (G-квадруплексы).
В четырехспиральные комплексы могут сплетаться четыре отдельные цепочки ДНК, но это скорее является исключением. Чаще единственная нить нуклеиновой кислоты просто завязывается в узел, образуя характерные утолщения (например, на концах хромосом), либо двуцепочечная ДНК на каком-то богатом гуанином участке образует локальный квадруплекс.
Наиболее изучено существование квадруплексов на концах хромосом - на теломерах и в онкопромоторах. Однако до сих пор полное представление о локализации такой ДНК в человеческих хромосомах не известно.
Все эти необычные структуры ДНК в линейной форме нестабильны по сравнению с В-формой ДНК. Однако ДНК часто существует в кольцевой форме топологического напряжения, когда у нее имеется так называемая сверхспирализация. В этих условиях легко образуются неканонические структуры ДНК: Z-формы, "кресты" и "шпильки", H-формы, гуаниновые квадруплексы и i-мотив.
- Суперспирализированная форма - отмечается при выделении из ядра клетки без повреждения пентозо-фосфатного остова. Имеет форму сверхскрученных замкнутых колец. В сверхскрученном состоянии двойная спираль ДНК хотя бы один раз "перекручена сама на себя", т. е. содержит хотя бы один супервиток (принимает форму восьмерки).
- Релаксированное состояние ДНК - наблюдается при одиночном разрыве (разрыве одной нити). При этом супервитки исчезают и ДНК принимает форму замкнутого кольца.
- Линейная форма ДНК - наблюдается при разрыве двух нитей двойной спирали.
Третичная структура ДНК
Третичная структура ДНК образуется в результате дополнительного скручивания в пространстве двуспиральной молекулы - ее суперспирализации. Суперспирализации молекулы ДНК в эукариотических клетках в отличие от прокариот осуществляется в форме комплексов с белками.
ДНК эукариот почти вся находится в хромосомах ядер, лишь небольшое количество ее содержится в митохондриях, а у растений и в пластидах. Основное вещество хромосом эукариотических клеток (в том числе и хромосом человека) - это хроматин, состоящий из двухцепочечной ДНК, гистоновых и негистоновых белков.
Гистоновые белки хроматина
Гистоны - простые белки, составляют до 50% хроматина. Во всех изученных клетках животных и растений обнаружено пять основных классов гистонов: H1, H2A, H2B, H3, H4, различающихся по размерам, аминокислотному составу и величине заряда (всегда положительный).
Гистон Н1 млекопитающих состоит из одной полипептидной цепи, содержащей примерно 215 аминокислот; размеры других же гистонов варьируют от 100 до 135 аминокислот. Все они спирализованы и скручены в глобулу диаметром около 2,5 нм, содержат необычно большое количество положительно заряженных аминокислот лизина и аргинина. Гистоны могут быть ацетилированы, метилированы, фосфорилированы, поли(АДФ)-рибозилированы, а гистоны Н2А и Н2В – ковалентно связаны с убиквитином. Какова роль таких модификаций в становлении структуры и выполнении функций гистонами до конца пока не выяснено. Предполагается, что в этом заключается их способность взаимодействовать с ДНК и обеспечивать один из механизмов регуляции действия генов.
Гистоны взаимодействуют с ДНК в основном через ионные связи (солевые мостики), образующиеся между отрицательно заряженными фосфатными группами ДНК и положительно заряженными лизиновыми и аргининовыми остатками гистонов.
Негистоновые белки хроматина
Негистоновые белки в отличие от гистонов очень разнообразны. Выделено до 590 разных фракций ДНК-связывающих негистоновых белков. Их еще называют кислыми белками, так как в их структуре преобладают кислые аминокислоты (они являются полианионами). С разнообразием негистоновых белков связывают специфическую регуляцию активности хроматина. Например ферменты, необходимые для репликации и экспрессии ДНК, могут связываться с хроматином временно. Другие белки, скажем, принимающие участие в различных процессах регуляции, связываются с ДНК только в специфических тканях или на определенных стадиях дифференциации. Каждый белок комплементарен определённой последовательности нуклеотидов ДНК (сайт ДНК). К этой группе относят:
- семейство сайт-специфических белков типа "цинковые пальцы". Каждый "цинковый палец" узнаёт определённый сайт, состоящий из 5 нуклеотидных пар.
- семейство сайт-специфических белков - гомодимеры. Фрагмент такого белка, контактирующий с ДНК, имеет структуру "спираль-поворот-спираль".
- белки высокой подвижности (HMG-белки - от англ, high mobility gel proteins) - группа структурных и регуляторных белков, которые постоянно ассоциированы с хроматином. Они имеют молекулярную массу менее 30 кД и характеризуются высоким содержанием заряженных аминокислот. Благодаря небольшой молекулярной массе HMG-белки обладают высокой подвижностью при электрофорезе в полиакриламидном геле.
- ферменты репликации, транскрипции и репарации.
При участии структурных, регуляторных белков и ферментов, участвующих в синтезе ДНК и РНК, нить нуклеосом преобразуется в высококонденсированный комплекс белков и ДНК. Образованная структура в 10 000 раз короче исходной молекулы ДНК.

Хроматин
Хроматин - это комплекс белков с ядерной ДНК и неорганическими веществами. Основная часть хроматина неактивна. Она содержит плотно упакованную, конденсированную ДНК. Это гетерохроматин. Различают конститутивный, генетически неактивный хроматин (сателлитная ДНК) состоящий из неэкспрессируемых областей, и факультативный - неактивный в ряду поколений, но при определенных обстоятельствах способный эспрессировать.
Активный хроматин (эухроматин) неконденсированный, т.е. упакован менее плотно. В разных клетках его содержание составляет от 2 до 11%. В клетках головного мозга его больше всего - 10-11%, в клетках печени - 3-4 и почек - 2-3%. Отмечается активная транскрипция эухроматина. При этом его структурная организация позволяет использовать одну и ту же генетическую информацию ДНК, присущую данному виду организма, по-разному в специализированных клетках.
В электронном микроскопе изображение хроматина напоминает бусы: шаровидные утолщения размером около 10 нм, разделенные нитевидными перемычками. Эти шаровидные утолщения названы нуклеосомами. Нуклеосома является структурной единицей хроматина. Каждая нуклеосома содержит сверхспиральный сегмент ДНК длиной 146 пар нуклеотидов, намотанный с образованием 1,75 левых витков на нуклеосомный кор. Нуклеосомный кор – это гистоновый октамер, состоящий из гистонов Н2А, Н2В, Н3 и Н4, по две молекулы каждого вида (рис. 9), который выглядит как диск диаметром 11 нм и толщиной 5,7 нм. Пятый гистон, Н1, не входит в состав нуклеосомного кора и не участвует в процессе наматывания ДНК на гистоновый октамер. Он контактирует с ДНК в тех местах, где двойная спираль входит и выходит из нуклеосомного кора. Это межкоровые (линкерные) участки ДНК, длина которых варьирует в зависимости от типа клеток от 40 до 50 нуклеотидных пар. В результате этого варьирует и длина фрагмента ДНК, входящего в состав нуклеосом (от 186 до 196 нуклеотидных пар).
В состав нуклеосом входит примерно 90% ДНК, остальная ее часть приходится на линкер. Считается, что нуклеосомы - это фрагменты "молчащего" хроматина, а линкер - активного. Однако нуклеосомы могут развертываться и переходить в линейную форму. Развернутые нуклеосомы являются уже активным хроматином. Так наглядно проявляется зависимость функции от структуры. Можно считать, что чем больше хроматина находится в составе глобулярных нуклеосом, тем менее он активен. Очевидно, в разных клетках неодинаковая доля покоящегося хроматина связана с количеством таких нуклеосом.
На электронно-микроскопических фотографиях в зависимости от условий выделения и степени растяжения хроматин может выглядеть не только как длинная нить с утолщениями – "бусинками" нуклеосом, но и как более короткая и более плотная фибрилла (волокно) диаметром 30 нм, образование которой наблюдается при взаимодействии гистона Н1, связанного с линкерным участком ДНК и гистона Н3, что приводит к дополнительному скручиванию спирали из шести нуклеосом на виток с образованием соленоида диаметром 30 нм. При этом гистоновый белок может препятствовать транскрипции ряда генов и таким образом регулировать их активность.
В результате описанных выше взаимодействий ДНК с гистонами сегмент двойной спирали ДНК из 186 пар оснований со средним диаметром 2 нм и длиной 57 нм превращается в спираль диаметром 10 нм и длиной 5 нм. При последующем сжатии этой спирали до волокна диаметром 30 нм степень конденсации увеличивается еще в шесть раз.
В конечном итоге упаковка дуплекса ДНК с пятью гистонами приводит к 50-кратной конденсации ДНК. Однако даже столь высокая степень конденсации не может объяснить почти 50 000 - 100 000-кратное уплотнение ДНК в метафазной хромосоме. К сожалению детали дальнейшей упаковки хроматина вплоть до метафазной хромосомы пока не известны, поэтому можно рассматривать лишь общие особенности этого процесса.
Уровни компактизации ДНК в хромосомах
Каждая молекула ДНК упакована в отдельную хромосому. В диплоидных клетках человека содержится 46 хромосом, которые располагаются в ядре клетки. Общая длина ДНК всех хромосом клетки составляет 1,74 м, однако диаметр ядра, в которое упакованы хромосомы, в миллионы раз меньше. Такая компактная укладка ДНК в хромосомах и хромосом в ядре клетки обеспечивается разнообразными, гистоновыми и негистоновыми белками, взаимодействующими в определенной последовательности с ДНК (см выше). Компактизация ДНК в хромосомах позволяет уменьшить ее линейные размеры примерно в 10 000 раз - условно с 5 см до 5 мкм. Выделяют несколько уровней компактизации (рис. 10).


- двойная спираль ДНК - отрицательно заряженная молекула диаметром 2 нм и длиной несколько см.
- нуклеосомный уровень
- хроматин выглядит в электронном микроскопе как цепочка "бусин" – нуклеосом - "на нити". Нуклеосома - это универсальная структурная
единица, которая обнаруживается как в эухроматине, так и в гетерохроматине, в интерфазном ядре и метафазных хромосомах.
Нуклеосомный уровень компактизации обеспечивается специальными белками - гистонами. Восемь положительно заряженных гистоновых доменов образуют кор (сердцевину) нуклеосомы на которую наматывается отрицательно заряженная молекула ДНК. Это дает укорочение в 7 раз, при этом диаметр увеличивается с 2 до 11 нм.
- соленоидный уровень
Соленоидный уровень организации хромосом характеризуется скручиванием нуклеосомной нити и образованием из нее более толстых фибрилл 20-35 нм в диаметре - соленоидов или супербидов. Шаг соленоида равен 11 нм, на один виток приходится около 6-10 нуклеосом. Соленоидная упаковка считается наиболее вероятной, чем супербидная, согласно которой фибрилла хроматина диаметром 20-35 нм представляет собой цепь гранул, или супербидов, каждая из которых состоит из восьми нуклеосом. На соленоидном уровне линейный размер ДНК сокращается в 6-10 раз, диаметр увеличивается до 30 нм.
- петлевой уровень
Петлевой уровень обеспечивается негистоновыми сайт-специфическими ДНК-связывающими белками, которые распознают определенные последовательности ДНК и связываются с ними, образуя петли примерно по 30-300 тысяч пар оснований. Петля обеспечивает экспрессию генов, т.е. петля является не только структурным, но и функциональным образованием. Укорочение на этом уровне происходит в 20-30 раз. Диаметр увеличивается до 300 нм. Петлеобразные структуры типа "ламповых щеток" в ооцитах земноводных можно видеть на цитологических препаратах. Эти петли, видимо, суперспирализованы и представляют собой домены ДНК, соответствующие, вероятно, единицам транскрипции и репликации хроматина. Специфические белки фиксируют основания петель и, возможно, некоторые их внутренние участки. Петлеобразная доменная организация способствует укладке хроматина в метафазных хромосомах в спиральные структуры более высоких порядков.
- доменный уровень
Доменный уровень организации хромосом изучен недостаточно. На данном уровне отмечается образование петлевых доменов - структур из нитей (фибрилл) толщиной 25-30 нм, которые содержат 60% белка, 35% ДНК и 5% РНК, практически не видны во всех фазах клеточного цикла за исключением митоза и несколько беспорядочно распределены по клеточному ядру. Петлеобразные структуры типа "ламповых щеток" в ооцитах земноводных можно видеть на цитологических препаратах.
Петлевые домены своим основанием прикрепляются к внутриядерному белковому матриксу в так называемых встроенных местах прикрепления, часто обозначаемых как MAR/SAR-последовательности (MAR, от англ. matrix associated region; SAR, от англ. scaffold attachment regions) – фрагментах ДНК протяженностью в несколько сотен пар оснований, которые характеризуются высоким содержанием (>65%) А/Т пар нуклеотидов. Каждый домен, по-видимому, имеет одну точку начала репликации и функционирует как автономная сверхспиральная единица. Любой петельный домен содержит множество единиц транскрипции, функционирование которых, вероятно, координируется – весь домен находиться либо в активном, либо в неактивном состоянии.
На доменном уровне в результате последовательной упаковки хроматина присходит уменьшение линейных размеров ДНК примерно в 200 раз (700 нм).
- хромосомный уровень
На хромосомном уровне происходит конденсация профазной хромосомы в метафазную с уплотнением петельных доменов вокруг осевого каркаса негистоновых белков. Эта суперспирализация сопровождается фосфорилированием в клетке всех молекул H1. В результате метафазную хромосому можно изобразить в виде плотно уложенных соленоидных петель, свернутых в тугую спираль. Типичная хромосома человека может содержать до 2600 петель. Толщина такой структуры достигает 1400 нм (две хроматиды), а молекула ДНК при этом укорачивается в 104 раз, т.е. с 5 см растянутой ДНК до 5 мкм.
Функции хромосом
Во взаимодействии с внехромосомными механизмами хромосомы обеспечивают
- хранение наследственной информации
- использование этой информации для создания и поддержания клеточной организации
- регуляцию считывания наследственной информации
- самоудвоение генетического материала
- передачу генетического материала от материнской клетки дочерним.
Существуют данные, что при активировании участка хроматина, т.е. при транскрипции, с него обратимо удаляются сначала гистон H1, а затем и октет гистонов. Это вызывает деконденсацию хроматина, последовательный переход 30-нанометровой фибриллы хроматина в 10-нанометровую нить и ее дальнейшее разворачивание в участки свободной ДНК, т.е. утрату нуклеосомной структуры.
Аббревиатура клеточный ДНК многим знакома из школьного курса биологии, но мало кто сможет с легкостью ответить, что это. Лишь смутное представление о наследственности и генетике остается в памяти сразу после окончания учебы. Знание, что такое ДНК, какое влияние оно оказывает на нашу жизнь, порой может оказаться очень нужным.
Молекула ДНК
Биохимики выделяют три типа макромолекул: ДНК, РНК и белки. Дезоксирибонуклеиновая кислота – это биополимер, который несет ответственность за передачу данных о наследственных чертах, особенностях и развитии вида из поколения в поколение. Его мономером является нуклеотид. Что такое молекулы ДНК? Это главный компонент хромосом и содержит генетический код.
Структура ДНК
Ранее ученые представляли, что модель строения ДНК периодическая, где повторяются одинаковые группы нуклеотидов (комбинаций молекул фосфата и сахара). Определенная комбинация последовательности нуклеотидов предоставляет возможность «кодировать» информацию. Благодаря исследованиям выяснилось, что у разных организмов структура различается.
Особенно известны в изучении вопроса, что такое ДНК американские ученые Александер Рич, Дэйвид Дэйвис и Гэри Фелзенфелд. Они в 1957 году представили описание нуклеиновой кислоты из трех спиралей. Спустя 28 лет, ученый Максим Давидович Франк-Каменицкий продемонстрировал, как дезоксирибонуклеиновая кислота, которая состоит из двух спиралей, складывается Н-образной формой из 3 нитей.
Структура у дезоксирибонуклеиновой кислоты двухцепочечная. В ней нуклеотиды попарно соединены в длинные полинуклеотидные цепи. Эти цепочки при помощи водородных связей делают возможным образование двойной спирали. Исключение – вирусы, у которых одноцепочечный геном. Существуют линейные ДНК (некоторые вирусы, бактерии) и кольцевые (митохондрии, хлоропласты).
Состав ДНК
Без знания, из чего состоит ДНК, не было бы ни одного достижения медицины. Каждый нуклеотид – это три части: остаток сахара пентозы, азотистое основание, остаток фосфорной кислоты. Исходя из особенностей соединения, кислоты могут называться дезоксирибонуклеиновой или рибонуклеиновой. В состав ДНК входит огромное число мононуклеотидов из двух оснований: цитозин и тимин. Кроме этого, она содержит производные пиримидинов, аденин и гуанин.
Есть в биологии определение DNA – мусорная ДНК. Функции ее еще неизвестны. Альтернативная версия названия – «некодирующая», что не верно, т.к. она содержит кодирующие белки, транспозоны, но их назначение тоже тайна. Одна из рабочих гипотез говорит о том, что некоторое количество этой макромолекулы способствует структурной стабилизации генома в отношении мутаций.
Где находится
Расположение внутри клетки зависит от особенностей вида. У одноклеточных ДНК находится в мембране. У остальных живых существ она располагается в ядре, пластидах и митохондриях. Если говорить о человеческой ДНК, то ее называют хромосомой. Правда, это не совсем так, ведь хромосомы – это комплекс хроматина и дезоксирибонуклеиновой кислоты.
Роль в клетке
Основная роль ДНК в клетках – передача наследственных генов и выживание будущего поколения. От нее зависят не только внешние данные будущей особи, но и ее характер и здоровье. Дезоксирибонуклеиновая кислота находится в суперскрученном состоянии, но для качественного процесса жизнедеятельности она должна быть раскрученной. С этим ей помогают ферменты - топоизомеразы и хеликазы.
Топоизомеразы относятся к нуклеазам, они способны изменять степень скрученности. Еще одна их функция – участие в транскрипции и репликации (делении клеток). Хеликазы разрывают водородные связи между основаниями. Существуют ферменты лигазы, которые нарушенные связи «сшивают», и полимеразы, которые участвуют в синтезе новых цепей полинуклеотидов.

Как расшифровывается ДНК
Эта аббревиатура для биологии является привычной. Полное название ДНК- дезоксирибонуклеиновая кислота. Произнести такое не каждому под силу с первого раза, поэтому часто в речи расшифровка ДНК опускается. Встречается еще понятие РНК – рибонуклеиновая кислота, которая состоит из последовательностей аминокислот в белках. Они напрямую связаны, а РНК является второй по важности макромолекулой.
ДНК человека
Человеческие хромосомы внутри ядра разделены, что делает ДНК человека самым стабильным, полным носителем информации. Во время генетической рекомбинации спирали разделяются, происходит обмен участками, а затем связь восстанавливается. За счет повреждения ДНК образовываются новые комбинации и рисунки. Весь механизм способствует естественному отбору. До сих пор неизвестно, как долго она отвечает за передачу генома, и какова ее эволюция метаболизма.
Кто открыл
Первое открытие структуры ДНК приписывают английским биологам Джеймсу Уотсону и Френсису Крику, которые в 1953 году раскрыли особенности строения молекулы. Нашел же ее в 1869 году швейцарский врач Фридрих Мишер. Он изучал химический состав животных клеток с помощью лейкоцитов, которые массово скапливаются в гнойных поражениях.
Мишер занимался изучением способов отмывания лейкоцитов, выделял белки, когда обнаружил, что кроме них есть что-то еще. На дне посуды во время обработки образовался осадок из хлопьев. Изучив эти отложения под микроскопом, молодой врач обнаружил ядра, которые оставались после обработки соляной кислотой. Там содержалось соединение, которое Фридрих назвал нуклеином (от лат. nucleus - ядро).
